Содержание
Является ли подсолнух зерновой культурой?
Перейти к основному содержанию
Skip to search
Вы здесь
- Главная »
- Разделы форума »
- Сад и огород, полеводство и земледелие »
- Общие вопросы по полеводству, земледелию, садоводству и огородничеству, вопросы по земле »
- Является ли подсолнух зерновой культурой?
На форуме
В блогах
- Эксперты озвучили схемы протравливания семян яровых на Агрополигоне «…
- Сколько стоит построить коровник
- Силос для свиней
- Записки пасечника: зимовка пчел
- Выгодный бизнес – выращивание тыквы на семена
В статьях
Сохранить материал к себе
Рекомендуемые статьи
Похожие материалы
- Люпин является непревзойденной одногодичной сидеральной культурой
- В Волгоградской области в 2020 году увеличена площадь посева масличных, основной культурой является подсолнечник
- В Воронежской области вырос подсолнух-гигант
- Создание зерновой биржи без участия ИДК является результатом интриг? – мнения
- Продам мед (разнотравье, подсолнух)
- Кукуруза станет второй экспортной культурой в России после пшеницы
- Закупаю масличный подсолнух
В видео
Буртование сапропеля |
Весна пришла, поход за вербой, обзор хозяйства |
Бентамские карликовые куры — лучшие несушки? |
Как отличить селезня от уточки у мускусных утят |
Содержание гусей на даче в СНТ |
Зима в деревне на Смоленщине |
Все видео
В объявлениях
| Авиахимобработка полей вертолетами | |
| Защита посевов от лугового мотылька и его гусеницы с вертолета Ми-2 самолета Ан-2 | |
| Внесение гербицидов агродронами дельтапланами самолетами малой авиации | |
| Весенняя подкормка озимых культур с вертолета | |
| Участок 56 соток берег Дона Азовский район | |
| Натуральный сушеный порошок, чипсы из овощей и грибов |
- Инкубация яиц
- Ветеринария
- Инвентарь и оборудование пчеловода
- Откорм скота
- Откармливаем бройлеров до осени
- Как вырастить дома саженцы винограда для весенней высадки
- Ветеринарные мероприятия для поросят в первые дни жизни
- Устройство задних мостов колесных тракторов
Активно обсуждаем
- Корова съела пакет с хозяйственным мылом
- Как найти сбыт на козье молоко?
- Кролик ходит на диван
- К цели полностью автономного хозяйства
- Ищу работу на постоянной основе!
Ещё
Я-фермер
Ваше мнение
В ЛПХ и КФХ
| ТК Авиком — полнорационные комбикорма и белково-витаминно-минеральные концентраты | |
| ФХ «Аделаида» | |
| АгроГород ООО | |
| ООО Экосистемы Мегаполиса — строительство объектов животноводства «под ключ» |
Главное меню
Перевозка подсолнечника: нюансы и тонкости
Подсолнечник — зерновая культура, значение которого для различных сфер экономики очень велико. В наших широтах это основная масличная культура. Семена подсолнечника содержат 50 — 52 % пищевого масла и 16 % белка. Поэтому профессиональная и грамотная перевозка подсолнечника необходима.
В наших широтах это основная масличная культура. Семена подсолнечника содержат 50 — 52 % пищевого масла и 16 % белка. Поэтому профессиональная и грамотная перевозка подсолнечника необходима.
К тому же, масло подсолнечника имеет высокие вкусовые качества и превосходит многие растительные жиры по питательности и усвояемости. Его используют непосредственно в пищу, а также при изготовлении маргарина, мучных изделий, консервов, кондитерки.
Нет смысла перечислять содержание полезных для организма человека полезных веществ.
Наличие в составе рациона питания человека этого продукта, ускоряют расщепление холестерина в организме, что положительно влияет на состояние здоровья.
Некоторые сорта масла подсолнечника служат сырьем для мыловаренной и лакокрасочной промышленности, применяются в производстве стеарина, клеенки, линолеума, электроарматуры, водонепроницаемых тканей, и пр.
Перерабатывая семена на масло, получают продукты и шрот, которые являются ценным высокобелковым кормом с протеином с большим количеством аминокислот. В 1 кг шрота содержится 1,02 корм. ед. и 363 г усваиваемого протеина, а в 1 кг жмыха – 1,09 корм. ед. и 226 г усваиваемого протеина.
В 1 кг шрота содержится 1,02 корм. ед. и 363 г усваиваемого протеина, а в 1 кг жмыха – 1,09 корм. ед. и 226 г усваиваемого протеина.
Лузга семян подсолнечника — ценное сырье при производстве гексозного и пентозного сахара, использующиеся при производстве этилового спирта и кормовых дрожжей и получения форфурола для производства пластмасс, небьющегося стекла, искусственного волокна и других химических материалов.
Виды транспортировки подсолнечника
Из вышесказанного можно становится ясно, насколько важна качественная перевозка семечки с поля к производителям масла и на элеваторы, на тока и специальные площадки, где хранится зерно.
Для транспортировки подсолнечника используются все виды транспорта — подвижной состав, автопоезда, специализированный автотранспорт, морской и авиатранспорт. Однако, к ним предъявляются специальные требования безопасной транспортировки.
На законодательном уровне специальными актами были установлены правила перевозки подсолнечника, комбикормов, жмыхов и шротов: в вагонах – зерновозах насыпью или навалом в упаковке в универсальном вагоне.
Используется абсолютно новый вид транспортировки — вагон — хоппер. В нем перевозят зерно и другие сыпучие пищевые продукты (сахар-сырец, подсолнечник, жом, отруби, шрот, комбикорм, и пр. которые требуют особенной защиты от атмосферных осадков). Вагон является принципиально новым видом доставки по железной дороге, он конкурирует на российском рынке грузоперевозок.
При транспортировке семян от железнодорожных станций до элеватора или масло перерабатывающего завода необходим специализированный автотранспорт. Продукт гигроскопичен, поэтому требует особых условий доставки. Современная технология транспортировки продукта — специальная тара, позволяющая безопасно, комфортно, удобно и эффективно производить операцию.
Специфика работ при перевозке семечек подсолнуха
Чтобы обеспечить рентабельность перевозки семечек, требуются грузовые автомобили, специально оборудованные под подсолнечник. Обычные зерновозы не всегда могут подойти для такого рода операций. Иногда требуется дополнительные затраты для их переоборудования.
Перевозка, семечки, зерновых, другой сельхозпродукции — важное и ответственное мероприятие. Ведь важно урожай убрать вовремя, а для этого приходится задействовать специализированный транспорт.
Первый способ транспортировки подсолнуха с поля — один из распространенных. Комбайн движется по полю и загружает рядом идущую специализированную машину, которая затем отвозит зерно на площадку временного хранения или элеватор. Это процесс сопряжен с большими расходами горючего и времени.
Можно семечки перевозить на площадку временного хранения, где ее очищают от мусора и грязи. А затем снова погрузчиком грузят на специализированный автотранспорт и везут на элеватор или завод по производству подсолнечного масла.
Также, с тока или площадки автотранспортом семечки перевозят к подвижному составу, который также, должен быть подготовлен для перевозки этого продукта.
Важно, чтобы перевозка семечки была эффективной и не могла испортить перевозимый груз, который отличается высокой гигроскопичностью.
Автодоставка
Что касается открытого автотранспорта грузоподъемностью от 20 до 35 тонн, то он в зависимости от того, какие возможности есть у погрузо-разгрузочных пунктов, может быть бортовым самосвалом или самосвальным автопоездом – зерновозом с задней и боковой выгрузкой, обязательно оснащенных брезентом или другой защитой от атмосферных осадков и ветра.
Автопоезда в сцепке с полуприцепами — это перевозка зерновых большой кубатурой. Особенно важно, что происходит более быстрая выгрузка. Так, разгрузка комплекса зерновоз — полуприцеп происходит в результате одного опрокидывания. Использование автосцепок с прицепами — это поэтапное взвешивание и без ограничений на приемных пунктах.
Использование зерна подсолнечника | Nuseed Europe
FacebookTwitter
Мировое производство семян подсолнечника неуклонно растет, прогнозируется, что урожай в текущем маркетинговом году составит около 54 миллионов метрических тонн.
На его долю приходится около 10% производства сырого растительного масла, что делает его третьей по важности масличной культурой после сои и рапса.
Украина, Россия, Европейский Союз и Аргентина являются основными странами-производителями семян, при этом на Украину и Россию приходится более половины мировых семян подсолнечника и почти 60% производства масла.
Использование семян подсолнечника
Масло
Крупнейшим сегментом рынка подсолнечника является извлечение масла путем дробления семян.
Затем масло используется в пищевой промышленности либо в качестве ингредиента для производства полуфабрикатов или готовых пищевых продуктов, либо оно разливается в бутылки в качестве конечного продукта.
Подсолнечное масло можно далее разделить на товарные масла, которые продаются в больших объемах и по низким ценам; и специальные масла, продаваемые в небольших объемах и по высоким ценам, такие как продукты премиум-класса и сертифицированные продукты, такие как органические и справедливая торговля.
Подсолнечное масло добывают тремя различными способами; холодный отжим; горячий пресс; и рафинированное подсолнечное масло.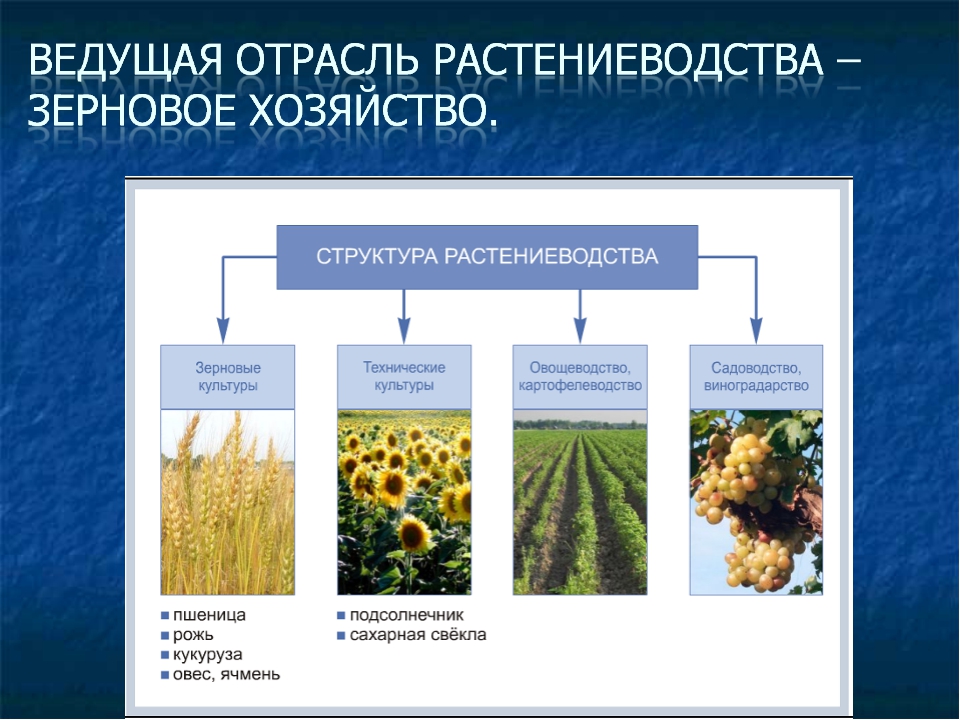
Существует небольшое, но растущее число кустарных переработчиков, производящих масла холодного отжима на фермах, которые продаются как ценный продукт, часто как органический и в сочетании с другими маслами и мукой.
Подсолнечное масло с высоким содержанием олеиновой кислоты предпочтительно для пищевой промышленности, поскольку оно естественно стабильно.
Еда
Мука, оставшаяся после извлечения масла, является богатым источником белка и отличным кормом для скота, особенно для жвачных животных.
По сравнению с соевым шрот подсолнечника содержит меньше энергии и аминокислоты лизина, но больше клетчатки и метионина.
Отделенная оболочка семян, или шелуха, содержит много клетчатки и мало белка, поэтому имеет низкую коммерческую кормовую ценность, поэтому ее часто сжигают в качестве источника тепла для дробильных установок.
На каждые 100 кг измельченных семян подсолнечника переработчик получает около 40 кг масла, 35 кг высокобелкового шрота и 25 кг побочных продуктов.
Здоровый спрос
Спрос на подсолнечное масло растет из года в год, в основном в результате повышения осведомленности о здоровье и признания преимуществ подсолнечного масла.
Подсолнечное масло богато витаминами А и Е, линолевой кислотой и мононасыщенными жирами, которые помогают контролировать уровень холестерина.
Кроме того, высокоолеиновые подсолнечные масла были модифицированы, чтобы быть более богатыми олеиновой кислотой, и производители продуктов питания обращаются к высокоолеиновым подсолнечным маслам в качестве замены вредных для здоровья трансжиров.
Подсолнечное масло светлого цвета и нейтрального вкуса является хорошим универсальным маслом, поскольку оно может выдерживать высокие температуры приготовления пищи.
Подсолнечное масло — одно из самых полезных растительных масел для приготовления пищи.
Кондитерские изделия
Семена подсолнечника также можно употреблять в пищу напрямую или с минимальной обработкой, и они называются кондитерскими подсолнухами.
В то время как подсолнечное масло и шрот производятся из одних и тех же сортов подсолнечника, семена для кондитерских изделий имеют свои особенности для их конкретных целей.
Как правило, семена кондитерских изделий полосатые, крупнее, чем масляные, с меньшим процентным содержанием масла и делятся на три категории.
В скорлупе
Это самые крупные семена, более 1,2 см в длину, с ядром, состоящим из половины веса, и толстой черно-белой полосатой оболочкой, продаются в скорлупе для перекуса.
Большинство из них обжариваются, иногда приправляются солью, а недавние разработки включают введение ароматизаторов, таких как барбекю, сметаны и лука.
Очищение от шелухи
Это семена среднего размера, механически очищенные от шелухи, ядро которых употребляют либо в качестве закуски, либо в качестве пищевого ингредиента в хлебобулочных изделиях.
Они широко используются в продуктах для здоровья и здоровых закусках, их можно добавлять в салаты, есть в чистом виде или измельчать до тонкой муки, а также использовать в качестве безглютеновой альтернативы муке с низким содержанием углеводов и орехов.
Корм для птиц
Это самые маленькие семена, которые продаются для кормления птиц или используются в кормах для домашних животных.
Мировой рынок кормов для птиц можно разделить на птиц, содержащихся в клетках, диких птиц, голубей (особенно гоночных голубей) и развивающийся рынок покровных культур, выращиваемых для охоты или создания среды обитания диких животных.
Возможности
Хотя точной информации о посевных площадях кондитерских сортов нет, ожидается, что кондитерские гибриды будут все чаще заменять обычные сорта, поскольку производители стремятся выйти на новые рынки.
По сравнению с масличными культурами, выращивание кондитерских подсолнухов более сложное, выбор подходящего сорта для выбранного вами рынка является ключевой отправной точкой, к счастью, у Nuseed есть ряд доступных гибридных кондитерских сортов [ссылка на продукты]
Производство высокоолеинового подсолнечного масла растет в Европе, но поставки остаются ограниченными, что указывает на будущие возможности для экспортеров из других стран.
Что ждет подсолнухи в будущем?
Существует ряд факторов, которые могут повлиять на использование подсолнечника в будущем, вот несколько ключевых моментов:
Здоровье
| Здоровая природа подсолнечного масла, вероятно, приведет к тому, что все больше производителей продуктов питания будут использовать его, поскольку они меняют свою продукцию, чтобы удовлетворить потребности более заботящихся о своем здоровье потребителей. |
| Высокоолеиновая | Обычные семена подсолнечника содержат 65% линолевой кислоты и 20% олеиновой кислоты, селекционеры вывели имеющиеся в продаже сорта подсолнечника с содержанием от 60% до 90% олеиновой кислоты. Это открыло возможности использования подсолнечного масла с высоким содержанием олеиновой кислоты в качестве стабилизатора пищевых продуктов, увеличения срока годности продуктов и в качестве полезного ингредиента. Страны Западной Европы переходят на высокоолеиновое подсолнечное масло, и производители выбирают высокоолеиновые сорта, чтобы воспользоваться этой рыночной возможностью. |
| Без глютена | Рост глютеновой болезни привел к сильному росту рынка безглютеновой муки, подсолнечная мука может использоваться в качестве безглютеновой альтернативы. |
| Будущие разработки | Подсолнечное масло в качестве заменителя арахисового масла в ситуациях, когда вызывает беспокойство аллергия на арахис. Корпуса, смешанные с клеем, сформованные в цилиндры, можно сжигать для обогрева, часто лишь с незначительными модификациями существующих котлов. |
| Без ГМО | Из-за высокого риска передачи генов от культивируемого подсолнечника к дикому маловероятно, что подсолнечник когда-либо получит разрешение на производство ГМО-сортов. |

Подсолнечник: потенциальная фруктановая культура?
Введение
Подсолнечник ( Helianthus annuus L. ) является важной сельскохозяйственной культурой во всем мире, занимая 4 th среди видов, производящих масло 1 , которые производят высококачественное масло для потребления человеком. У этого вида наполнение зерна в основном зависит от фактического фотосинтеза. Согласно подробному анализу С-бюджета (Hall et al., 1990), С, зафиксированный в период до цветения, составляет всего около 15% от общего поглощения С зерном на участках без дефицита воды. Повышение способности тканей стебля к накоплению запасов может способствовать увеличению наполнения зерна и особенно стабильности урожая. Как правило, зерна, расположенные во внутренней части головки (то есть самые молодые), не развиваются должным образом, оставляя центр заготовленного органа с большинством тщетных зерен. Известно, что это явление зависит от доступности фотоассимилятов (Alkio et al., 2003; Cantagallo et al., 2004), поскольку дефицит сосудистых связей не препятствует наполнению семян подсолнечника (Alkio and Grimm, 2003).
) является важной сельскохозяйственной культурой во всем мире, занимая 4 th среди видов, производящих масло 1 , которые производят высококачественное масло для потребления человеком. У этого вида наполнение зерна в основном зависит от фактического фотосинтеза. Согласно подробному анализу С-бюджета (Hall et al., 1990), С, зафиксированный в период до цветения, составляет всего около 15% от общего поглощения С зерном на участках без дефицита воды. Повышение способности тканей стебля к накоплению запасов может способствовать увеличению наполнения зерна и особенно стабильности урожая. Как правило, зерна, расположенные во внутренней части головки (то есть самые молодые), не развиваются должным образом, оставляя центр заготовленного органа с большинством тщетных зерен. Известно, что это явление зависит от доступности фотоассимилятов (Alkio et al., 2003; Cantagallo et al., 2004), поскольку дефицит сосудистых связей не препятствует наполнению семян подсолнечника (Alkio and Grimm, 2003). На самом деле, ситуация часто усугубляется возникновением засушливых периодов во время налива зерна, которые характерны для одного из основных регионов выращивания подсолнечника в мире, таких как аргентинские пампасы (Садрас и Холл, 19).89). В этих условиях скорость фотосинтеза растений подсолнечника может сильно снижаться. Холл и др. (1989, 1990) сообщили, что С, зафиксированный в период перед цветением, составляет около 27% от общего поглощения С зерном в культурах, подверженных водному стрессу. Несмотря на большую ремобилизацию углерода, в период водного стресса этого может быть недостаточно для компенсации нарушенной фиксации углерода, поскольку также затрагиваются физиологические процессы, такие как фотосинтез и рост. Таким образом, в засушливые годы часто наблюдаются серьезные потери урожая (Садрас и др., 19).93). Также ожидается снижение качества зерна, поскольку как относительное содержание масла, так и состав жирных кислот зависят от перехваченной ФАР (фотосинтетически активной радиации) во время налива зерна (Dosio et al.
На самом деле, ситуация часто усугубляется возникновением засушливых периодов во время налива зерна, которые характерны для одного из основных регионов выращивания подсолнечника в мире, таких как аргентинские пампасы (Садрас и Холл, 19).89). В этих условиях скорость фотосинтеза растений подсолнечника может сильно снижаться. Холл и др. (1989, 1990) сообщили, что С, зафиксированный в период перед цветением, составляет около 27% от общего поглощения С зерном в культурах, подверженных водному стрессу. Несмотря на большую ремобилизацию углерода, в период водного стресса этого может быть недостаточно для компенсации нарушенной фиксации углерода, поскольку также затрагиваются физиологические процессы, такие как фотосинтез и рост. Таким образом, в засушливые годы часто наблюдаются серьезные потери урожая (Садрас и др., 19).93). Также ожидается снижение качества зерна, поскольку как относительное содержание масла, так и состав жирных кислот зависят от перехваченной ФАР (фотосинтетически активной радиации) во время налива зерна (Dosio et al. , 2000; Izquierdo et al., 2009).
, 2000; Izquierdo et al., 2009).
Запасы сахара в стеблях подсолнечника состоят из сахарозы и моносахаридов, которые вместе с ограниченным количеством крахмала составляют запасы, которые могут быть ремобилизованы в зерне (Goldschmidt and Huber, 1992; Sims et al., 1999). Поскольку запасание С в виде моно- и дисахаридов предполагает высокое повышение осмоляльности клеток, накопление этих сахаров в стеблях приводит к быстрому увеличению объема клеток и тканей, что проявляется в основном по утолщению стеблей. Таким образом, часть избытка углерода направляется на вложение в структурные компоненты вместо того, чтобы оставаться в качестве резервных углеводов, которые можно было бы ремобилизовать в головной мозг. Полимеризация сахарозы во фруктаны могла бы дать возможность хранить гораздо большее количество ассимилятов на единиц объема выпуска без значительного осмотического эффекта. Соответственно, вводя гены фруктозилтрансферазы в растения сахарного тростника, Нелл (2007) обнаружил, что содержание сахара на 63% выше, чем в контрольных, нетрансформированных растениях. Сохраненные фруктаны, в свою очередь, при необходимости легко мобилизуются. У большинства мелкозерновых культур фруктаны запасаются в стебле и в дальнейшем способствуют наполнению зерна в большей степени, чем сахароза в подсолнечнике. Например, Боррелл и др. (1989) сообщили, что запасы стеблей после цветения могли составлять не менее 21% конечного урожая зерна полукарликовой пшеницы, в то время как Gebbing et al. (1999) сообщил о сходных средних значениях для двух сортов этого вида. Эти проценты увеличиваются в периоды засухи (Blum, 1998; Gupta et al., 2011). Поскольку резервные ассимиляты, хранящиеся в вегетативных органах растения до цветения, могут быть мобилизованы во время налива зерна, они могут защищать урожай зерна от неблагоприятных условий для фотосинтеза в этот период. Это общее явление наблюдается для зерновых (Gebbing et al., 1999) и других культур, включая подсолнечник (Hall et al., 19).89). Помимо предполагаемого влияния на накопление углерода и ремобилизацию в репродуктивных структурах, вероятность того, что подсолнечник накапливает фруктан, также может иметь прямое влияние на устойчивость сельскохозяйственных культур к летней засухе.
Сохраненные фруктаны, в свою очередь, при необходимости легко мобилизуются. У большинства мелкозерновых культур фруктаны запасаются в стебле и в дальнейшем способствуют наполнению зерна в большей степени, чем сахароза в подсолнечнике. Например, Боррелл и др. (1989) сообщили, что запасы стеблей после цветения могли составлять не менее 21% конечного урожая зерна полукарликовой пшеницы, в то время как Gebbing et al. (1999) сообщил о сходных средних значениях для двух сортов этого вида. Эти проценты увеличиваются в периоды засухи (Blum, 1998; Gupta et al., 2011). Поскольку резервные ассимиляты, хранящиеся в вегетативных органах растения до цветения, могут быть мобилизованы во время налива зерна, они могут защищать урожай зерна от неблагоприятных условий для фотосинтеза в этот период. Это общее явление наблюдается для зерновых (Gebbing et al., 1999) и других культур, включая подсолнечник (Hall et al., 19).89). Помимо предполагаемого влияния на накопление углерода и ремобилизацию в репродуктивных структурах, вероятность того, что подсолнечник накапливает фруктан, также может иметь прямое влияние на устойчивость сельскохозяйственных культур к летней засухе. Было продемонстрировано, что способность подсолнечника к осморегуляции тесно связана с поддержанием урожайности в засушливые годы (Chimenti et al., 2002). Известно, что фруктаны обеспечивают осмотическую регуляцию растений, поскольку присутствуют в виде континуума олигомеров, отличающихся друг от друга одним остатком фруктозы, и легко деполимеризуются в гексозы. Эта особенность позволяет растениям, содержащим фруктаны, извлекать воду при более низком водном потенциале почвы (Garcia et al., 2011). Более того, известно, что фруктаны защищают мембраны путем непосредственного внедрения в липиды и, таким образом, облегчают растениям стресс от засухи (Valluru and Van den Ende, 2008; Livingston et al., 2009).; Ван ден Энде, 2013).
Было продемонстрировано, что способность подсолнечника к осморегуляции тесно связана с поддержанием урожайности в засушливые годы (Chimenti et al., 2002). Известно, что фруктаны обеспечивают осмотическую регуляцию растений, поскольку присутствуют в виде континуума олигомеров, отличающихся друг от друга одним остатком фруктозы, и легко деполимеризуются в гексозы. Эта особенность позволяет растениям, содержащим фруктаны, извлекать воду при более низком водном потенциале почвы (Garcia et al., 2011). Более того, известно, что фруктаны защищают мембраны путем непосредственного внедрения в липиды и, таким образом, облегчают растениям стресс от засухи (Valluru and Van den Ende, 2008; Livingston et al., 2009).; Ван ден Энде, 2013).
В течение последних двух десятилетий было предпринято несколько попыток перенести гены фруктана от видов, несущих фруктан, на другие растения. В результате в нескольких случаях сообщалось об улучшении устойчивости к засухе или морозам (Pilon-Smits et al., 1995; Konstantinova et al. , 2002; Hisano et al., 2004; Parvanova et al., 2004; Bie et al. , 2012), но, как правило, успех таких трансформаций был довольно ограниченным, поскольку трансгенные растения склонны продуцировать фруктан в низкой концентрации (Cairns, 2003). Кроме того, в отношении подсолнечника эта возможность сомнительна, поскольку этот вид считался устойчивым к генетической трансформации из-за трудностей в процедурах регенерации растений (Moschen et al., 2014).
, 2002; Hisano et al., 2004; Parvanova et al., 2004; Bie et al. , 2012), но, как правило, успех таких трансформаций был довольно ограниченным, поскольку трансгенные растения склонны продуцировать фруктан в низкой концентрации (Cairns, 2003). Кроме того, в отношении подсолнечника эта возможность сомнительна, поскольку этот вид считался устойчивым к генетической трансформации из-за трудностей в процедурах регенерации растений (Moschen et al., 2014).
Подсолнечник является близким родственником топинамбура ( H. tuberosus L.), который накапливает большое количество фруктана инулинового типа в клубнях (которые представляют собой видоизмененные стебли), а также в настоящих стеблях и даже корнях (Seiler, 2007). На самом деле топинамбур (как сахарную культуру) можно выращивать как на клубни, так и на надземную биомассу, для которой существуют сорта, характеризующиеся хорошо развитыми стеблями и низкой урожайностью клубней (Caserta and Cervigni, 1991). Причина, по которой подсолнечник не накапливает фруктаны, неясна. В начале 20 — гг. М. Х. Колин во Франции провел эксперименты по прививке подсолнечника и топинамбура, которые выявили, что только части, принадлежащие последнему, накапливают инулин, независимо от того, какой вид находился в основании прививки (Колин, 1922). Первая возможность объяснить отсутствие фруктана в подсолнечнике заключается в том, что гены ферментов синтеза фруктана могут отсутствовать или не функционировать у этого вида. Вторая возможность заключается в том, что, даже если она присутствует, экспрессия ферментов, синтезирующих фруктан, в подсолнечнике подавляется из-за метаболических причин. Обе возможности проанализированы ниже.
В начале 20 — гг. М. Х. Колин во Франции провел эксперименты по прививке подсолнечника и топинамбура, которые выявили, что только части, принадлежащие последнему, накапливают инулин, независимо от того, какой вид находился в основании прививки (Колин, 1922). Первая возможность объяснить отсутствие фруктана в подсолнечнике заключается в том, что гены ферментов синтеза фруктана могут отсутствовать или не функционировать у этого вида. Вторая возможность заключается в том, что, даже если она присутствует, экспрессия ферментов, синтезирующих фруктан, в подсолнечнике подавляется из-за метаболических причин. Обе возможности проанализированы ниже.
Наличие ферментов синтеза фруктана в подсолнечнике
Исчерпывающий анализ возможности того, что гены ферментов синтеза фруктана в подсолнечнике могут отсутствовать или быть нефункциональными, еще не проводился. Это может быть связано с тем, что геном подсолнечника до сих пор недоступен. Принятая модель синтеза фруктана у высших растений предполагает наличие различных ферментов, в основном в том числе 1-SST (1-сахароза:сахарозофруктозилтрансфераза) и 1-FFT (фруктан:фруктан-1-фруктозилтрансфераза) для инициации и полимеризации фруктана соответственно (Vijn и Смикенс, 19 лет99). С целью выяснить, кодирует ли геном подсолнечника ферменты, синтезирующие фруктан, мы провели поиск гомологичных последовательностей 1-SST и 1-FFT из разных видов растений в общедоступной базе данных EST подсолнечника 2 (Отдел биоинформатики в INTA, Херлингем, Буэнос-Айрес, Аргентина). ). Из нескольких идентифицированных контигов, которые показали высокое сходство с инвертазами (INV) и фруктозилтрансферазами из генов нескольких видов растений, два показали высокое сходство с генами ферментов синтеза фруктана: HeAn_C_8450_Contig8450 (Hacontig8450) с 1-SST и HeAn_C_1289.4_Contig12894 (Ha12894) в 1-БПФ (таблица 1). Ранее было описано, что о развитии способности к трансфруктозилированию в INV свидетельствуют различные аминокислотные замены, например, W на Y или F и/или N на S в мотиве WMNDPNG и WGW на WGY или WGF (Lammens et al., 2012). В то время как большинство обнаруженных нами контигов, по-видимому, принадлежат к семейству INV (например, HeAn_C_12706_Contig12706, Ha12706), в Hacontig8450 и Ha12894 отсутствуют все незаменимые аминокислоты, которые характеризуют ферменты INV (таблица 1), что позволяет предположить, что они принадлежат к группе фруктозилтрансфераз.
С целью выяснить, кодирует ли геном подсолнечника ферменты, синтезирующие фруктан, мы провели поиск гомологичных последовательностей 1-SST и 1-FFT из разных видов растений в общедоступной базе данных EST подсолнечника 2 (Отдел биоинформатики в INTA, Херлингем, Буэнос-Айрес, Аргентина). ). Из нескольких идентифицированных контигов, которые показали высокое сходство с инвертазами (INV) и фруктозилтрансферазами из генов нескольких видов растений, два показали высокое сходство с генами ферментов синтеза фруктана: HeAn_C_8450_Contig8450 (Hacontig8450) с 1-SST и HeAn_C_1289.4_Contig12894 (Ha12894) в 1-БПФ (таблица 1). Ранее было описано, что о развитии способности к трансфруктозилированию в INV свидетельствуют различные аминокислотные замены, например, W на Y или F и/или N на S в мотиве WMNDPNG и WGW на WGY или WGF (Lammens et al., 2012). В то время как большинство обнаруженных нами контигов, по-видимому, принадлежат к семейству INV (например, HeAn_C_12706_Contig12706, Ha12706), в Hacontig8450 и Ha12894 отсутствуют все незаменимые аминокислоты, которые характеризуют ферменты INV (таблица 1), что позволяет предположить, что они принадлежат к группе фруктозилтрансфераз.
Таблица 1 в непосредственной близости от активного центра.
Регулирование ферментов, синтезирующих фруктан, в
Helianthus
Наиболее известная модель индукции синтеза фруктана была разработана на основе исследований Poaceae, таких как пшеница и ячмень. Согласно модели, сахароза должна превышать определенный порог, чтобы вызвать экспрессию ферментов, синтезирующих фруктан (Pollock et al., 2003). Следовательно, сахароза, по-видимому, играет двойную роль в метаболизме фруктана: она является одновременно основным субстратом, используемым в синтезе фруктана, и также запускает путь передачи сигнала, который индуцирует ферменты, синтезирующие фруктозил-сахарозу (Tognetti et al., 2013). Возможно, что в случае с подсолнечником накапливается недостаточное количество сахарозы для индукции 1-SST, либо вследствие более высокого порога индукции сахарозой у подсолнечника, чем у топинамбура, либо вследствие высокой активности фруктангидролазы. что может препятствовать накоплению фруктана в тканях подсолнечника.
Затем мы провели эксперимент, в котором уровни эндогенной сахарозы в стеблях подсолнечника были резко повышены путем удаления основного акцепторного органа (головки) для анализа возможной индукции синтеза фруктана. Коммерческий гибрид подсолнечника (VDh581, Advanta Semillas SAIC) был выращен в поле в Балькарсе, Аргентина, в потенциальных условиях для этой среды. Когда урожай достиг фенологической стадии Шнайтера и Миллера R3 (примерно за 120° дней до цветения при базовой температуре 6°С), половину растений обезглавливали. Эта обработка привела как к значительному (152%) увеличению диаметра стебля, так и к увеличению (более чем в 24 раза) общей концентрации сахара в тканях стебля по сравнению с интактными растениями (рис. 1А, В). Концентрация сахарозы в стебле обезглавленных растений колебалась от 5,73 до 7,45 мг сахарозы г 9 .0191 -1 FW, в зависимости от даты отбора проб. Таким образом, можно предположить, что сильное расширение ткани, вызванное декапитацией, препятствует достижению концентраций сахарозы, которые могли бы превысить 15 мг г -1 сырой массы тела, если бы объем стебля оставался неизменным. Сообщалось о пороговых значениях сахарозы для индукции фруктана у видов трав, близких к 15 мг г -1 сырой массы (Королева и др., 1998), хотя прямое сравнение между видами невозможно из-за изменения объема стебля подсолнечника. ВЭЖХ-анализ состава сахаров выявил присутствие 1-кестозы в стебле контрольных растений, но ее уровни не повышались при декапитации головок, несмотря на большое накопление сахарозы (рис. 1С), достигающее 0,24 мг/г -1 ФВ. С другой стороны, были обнаружены незначительные количества олигомеров с более высокой степенью полимеризации (не показаны).
Сообщалось о пороговых значениях сахарозы для индукции фруктана у видов трав, близких к 15 мг г -1 сырой массы (Королева и др., 1998), хотя прямое сравнение между видами невозможно из-за изменения объема стебля подсолнечника. ВЭЖХ-анализ состава сахаров выявил присутствие 1-кестозы в стебле контрольных растений, но ее уровни не повышались при декапитации головок, несмотря на большое накопление сахарозы (рис. 1С), достигающее 0,24 мг/г -1 ФВ. С другой стороны, были обнаружены незначительные количества олигомеров с более высокой степенью полимеризации (не показаны).
РИСУНОК 1. Накопление сахара и профили в стеблях подсолнечника контрольных и декапитированных растений. (A) Вид обезглавленного растения (слева), показывающий большое утолщение стебля и общую стимуляцию роста по сравнению с контрольным растением (справа). (B) Концентрация общих растворимых углеводов (TSC) в стеблях декапитированных (треугольники, пунктирная линия) и контрольных (кружки, сплошная линия) растений (±SE), измеренная спектрофотометрически из водных экстрактов по фенол-серному методу ( стрелка указывает дату отбора проб). (C) Хроматограммы ВЭЖХ водных экстрактов стеблей, разделенных на колонке Prevail carbs ES и обнаруженных с помощью детектора показателя преломления (RID).
(C) Хроматограммы ВЭЖХ водных экстрактов стеблей, разделенных на колонке Prevail carbs ES и обнаруженных с помощью детектора показателя преломления (RID).
Неспособность подсолнечника существенно накапливать фруктан, вероятно, не связана с высокой активностью фруктангидролазы, поскольку известно, что последний фермент ингибируется сахарозой у большинства видов (Van Riet et al., 2006). С другой стороны, возможно, что у подсолнечника (а может быть, и у сложноцветных вообще) одной сахарозы недостаточно для индукции синтеза фруктанов, в отличие от однодольных растений. В этом смысле, работая с культурами волосатых корней цикория, Kusch et al. (2009 г.) показали, что в дополнение к высоким уровням сахарозы для индукции ферментов, синтезирующих фруктан, требуются низкие уровни азота. Кроме того, из работы H. tuberosus представляется возможным, что инициация синтеза фруктана может быть связана с индукцией клубнеобразования, поскольку до начала роста клубней, по-видимому, не происходит полимеризации сахарозы (Monti et al. , 2005). Начало образования клубней регулируется гормонально, хорошо известным промотором является жасмоновая кислота (Koda et al., 1994). Углубление наших знаний о H. tuberosus биологии с такими исследованиями, как Jung et al. (2014) также может помочь найти ответы.
, 2005). Начало образования клубней регулируется гормонально, хорошо известным промотором является жасмоновая кислота (Koda et al., 1994). Углубление наших знаний о H. tuberosus биологии с такими исследованиями, как Jung et al. (2014) также может помочь найти ответы.
Заключение и перспективы на будущее
Наши результаты показывают, что неспособность подсолнечника существенно накапливать фруктан не связана ни с отсутствием функциональных генов, кодирующих ферменты синтеза фруктана, ни с недостаточным уровнем сахарозы для индукции синтезирующих фруктан ферментов в стеблях. Представлены данные, свидетельствующие о том, что основные синтезирующие фруктаны ферменты (1-SST и 1-FFT) действительно экспрессируются в подсолнечнике и что 1-кестоза (то есть продукт активности SST) также обнаруживается среди сахаров, хранящихся в стебле. Однако количество фруктана, по-видимому, незначительно, и его накопление, вероятно, не вызывается обработкой, повышающей концентрацию сахарозы в стебле. Эти данные свидетельствуют о том, что подсолнечник потенциально может представлять собой культуру, содержащую фруктаны, что может иметь важное значение с агрономической точки зрения. Повышение урожайности зерна и масличности, более удобный состав жирных кислот и особенно улучшение устойчивости урожая к засухе и другим абиотическим стрессам гипотетически можно было бы достичь, если бы в стебле подсолнечника удалось накопить значительное количество фруктанов. Тем не менее, необходимы большие усилия, чтобы выяснить, как эффективно проявляется способность подсолнечника накапливать фруктаны. Тщательное изучение регуляторных механизмов, участвующих в его синтезе, может быть первым шагом к достижению этой цели. 9 http://atgc-sur.inta.gob.ar/welcome/default/index
Эти данные свидетельствуют о том, что подсолнечник потенциально может представлять собой культуру, содержащую фруктаны, что может иметь важное значение с агрономической точки зрения. Повышение урожайности зерна и масличности, более удобный состав жирных кислот и особенно улучшение устойчивости урожая к засухе и другим абиотическим стрессам гипотетически можно было бы достичь, если бы в стебле подсолнечника удалось накопить значительное количество фруктанов. Тем не менее, необходимы большие усилия, чтобы выяснить, как эффективно проявляется способность подсолнечника накапливать фруктаны. Тщательное изучение регуляторных механизмов, участвующих в его синтезе, может быть первым шагом к достижению этой цели. 9 http://atgc-sur.inta.gob.ar/welcome/default/index
Ссылки
Алкио М. и Гримм Э. (2003). Сосудистые связи между цветоложем и пустыми семянками подсолнечника ( Helianthus annuus L.). Дж. Экспл. Бот. 54, 345–348. doi: 10.1093/jxb/erg019
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Алкио М. , Шуберт А., Дипенброк В. и Гримм Э. (2003). Влияние коэффициента сток-источник на завязываемость и наполнение семян подсолнечника ( Helianthus annuus L.). Окружающая среда растительных клеток. 26, 1609–1619. doi: 10.1046/j.0016-8025.2003.01077.x
, Шуберт А., Дипенброк В. и Гримм Э. (2003). Влияние коэффициента сток-источник на завязываемость и наполнение семян подсолнечника ( Helianthus annuus L.). Окружающая среда растительных клеток. 26, 1609–1619. doi: 10.1046/j.0016-8025.2003.01077.x
Полный текст CrossRef | Google Scholar
Bie, X.L., Wang, K., She, M., Du, L., Zhang, S., Li, J., et al. (2012). Комбинированная трансформация трех генов пшеницы, кодирующих ферменты биосинтеза фруктана, обеспечивает повышенное содержание фруктана и устойчивость табака к абиотическим стрессам. Представитель клеток растений 31, 2229–2238. doi: 10.1007/s00299-012-1332-y
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Блюм, А. (1998). Улучшение наполнения зерна пшеницы в условиях стресса за счет мобилизации стеблевого резерва. Euphytica 100, 77–83. doi: 10.1023/A:1018303922482
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Боррелл А. К., Инколл Л.Д., Симпсон Р.Дж. и Даллинг М.Дж. (1989). Распределение сухого вещества и отложение и использование запасов стебля в посевах полукарликовой пшеницы. Энн. Бот. 63, 527–539.
К., Инколл Л.Д., Симпсон Р.Дж. и Даллинг М.Дж. (1989). Распределение сухого вещества и отложение и использование запасов стебля в посевах полукарликовой пшеницы. Энн. Бот. 63, 527–539.
Google Scholar
Кэрнс, А. Дж. (2003). Биосинтез фруктана в трансгенных растениях. Дж. Экспл. Бот. 54, 549–567. doi: 10.1093/jxb/erg056
Полный текст CrossRef | Google Scholar
Кантагалло, Дж. Э., Медан, Д., и Холл, А. Дж. (2004). Количество зерен подсолнечника в зависимости от затенения во время роста цветков, цветения и завязывания зерна. Полевые культуры Res. 85, 191–202. doi: 10.1016/S0378-4290(03)00160-6
Полный текст CrossRef | Академия Google
Казерта Г. и Червиньи Т. (1991). Использование стеблей топинамбура для производства фруктозы или этанола. Биоресурс. Технол. 35, 247–250. doi: 10.1016/0960-8524(91)
-YПолный текст CrossRef | Google Scholar
Чименти, К.А., Пирсон, Дж., и Холл, А.Дж. (2002). Осмотическая адаптация и сохранение урожая подсолнечника в условиях засухи. Полевые культуры Res. 75, 235–246. doi: 10.1016/S0378-4290(02)00029-1
Осмотическая адаптация и сохранение урожая подсолнечника в условиях засухи. Полевые культуры Res. 75, 235–246. doi: 10.1016/S0378-4290(02)00029-1
Полный текст CrossRef | Академия Google
Колин, Х. (1922). L’inuline dans les plantes greffées. La greffe soleil annuel topinambour. Преподобный генерал. Бот. 34, 202–213.
Google Scholar
Досио, Г.А.А., Агирресабаль, Л.А.Н., Андраде, Ф.Х., и Перейра, В.Р. (2000). Солнечное излучение, перехваченное при наливе семян и производстве масла у двух гибридов подсолнечника. Растениеводство. 40, 1637–1644. doi: 10.2135/cropsci2000.4061637x
Полный текст CrossRef | Google Scholar
Гарсия П.М.А., Асега А.Ф., Сильва Э.А. и Карвалью М.А.М. (2011). Влияние засухи и повторного полива на метаболизм фруктана в Vernonia herbaceae (Vell.) Rusby. Завод физиол. Биохим. 49, 664–670. doi: 10.1016/j.plaphy.2011.03.014
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Геббинг Т. , Шнайдер Х. и Кюбаух В. (1999). Использование предцветовых резервов при наливе зерна пшеницы. Оценка путем мечения устойчивых 13CO2/12CO2. Окружающая среда растительных клеток. 22, 851–858. doi: 10.1046/j.1365-3040.1999.00436.x
, Шнайдер Х. и Кюбаух В. (1999). Использование предцветовых резервов при наливе зерна пшеницы. Оценка путем мечения устойчивых 13CO2/12CO2. Окружающая среда растительных клеток. 22, 851–858. doi: 10.1046/j.1365-3040.1999.00436.x
Полный текст CrossRef | Академия Google
Goldschmidt, E.E., and Huber, S.C. (1992). Регуляция фотосинтеза путем накопления конечных продуктов в листьях растений, запасающих крахмал, сахарозу и гексозные сахара. Завод физиол. 99, 1443–1448. doi: 10.1104/pp.99.4.1443
CrossRef Full Text | Google Scholar
Гупта А.К., Каур К. и Каур Н. (2011). Мобилизация стеблевого резерва и акцепторная активность пшеницы в условиях засухи. утра. Дж. Растениевод. 2, 70–77. doi: 10.4236/ajps.2011.21010
Полнотекстовая перекрестная ссылка | Google Scholar
Холл, А. Дж., Коннор, Д. Дж., и Уитфилд, Д. М. (1989). Вклад предцветниковых ассимилятов в налив зерна у орошаемых и вододефицитных посевов подсолнечника I. Оценки по меченому углероду. Полевые культуры Res. 20, 95–112. doi: 10.1016/0378-4290(89)
Оценки по меченому углероду. Полевые культуры Res. 20, 95–112. doi: 10.1016/0378-4290(89)
-5
CrossRef Full Text | Google Scholar
Холл, А. Дж., Уитфилд, Д. М., и Коннор, Д. Дж. (1990). Вклад предцветниковых ассимилятов в налив зерна орошаемых и вододефицитных посевов подсолнечника II. Оценки из углеродного бюджета. Полевые культуры Res. 24, 273–294. doi: 10.1016/0378-4290(90)
-C
Полный текст CrossRef | Google Scholar
Хисано Х., Канадзава А., Каваками А., Ёсида М., Симамото Ю. и Ямада Т. (2004). Трансгенные растения райграса пастбищного, экспрессирующие гены фруктозилтрансфераз пшеницы, накапливают повышенное количество фруктана и приобретают повышенную устойчивость на клеточном уровне к замораживанию. Растениевод. 167, 861–868. doi: 10.1016/j.plantsci.2004.05.037
Полный текст CrossRef | Академия Google
Искьердо, Н.Г., Агирресабаль, Л.А.Н., Андраде, Ф.Х., Герудет, К., Валентинуз, О., и Перейра Ираола, М. (2009). Перехваченная солнечная радиация влияет на состав жирных кислот в масле сельскохозяйственных культур. Полевые культуры Res. 114, 66–74. doi: 10.1016/j.fcr.2009.07.007
(2009). Перехваченная солнечная радиация влияет на состав жирных кислот в масле сельскохозяйственных культур. Полевые культуры Res. 114, 66–74. doi: 10.1016/j.fcr.2009.07.007
Полный текст CrossRef | Google Scholar
Jung, W.Y., Lee, S.S., Kim, C.W., Kim, H.-S., Min, S.R., Moon, J.S., et al. (2014). Анализ RNA-Seq и сборка транскриптома de novo топинамбура ( Helianthus tuberosus Linne). PLoS ONE 9:e111982. doi: 10.1371/journal.pone.0111982
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кода Ю., Такахаши К. и Кикута Ю. (1994). Участие жасмоновой кислоты и родственных соединений в клубнеобразовании растений топинамбура ( Helianthus tuberosus L.). япон. J. Crop Sci. 63, 333–338. doi: 10.1626/jcs.63.3331626
Полный текст CrossRef | Академия Google
Константинова Т., Пырванова Д., Атанасов А. и Джилианов Д. (2002). Устойчивый к замерзанию табак, преобразованный для накопления осмопротекторов. Растениевод. 163, 157–164. doi: 10.1016/S0168-9452(02)00090-0
Растениевод. 163, 157–164. doi: 10.1016/S0168-9452(02)00090-0
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Королева О.А., Фаррар Дж.Ф., Дери Томос А. и Поллок С.Дж. (1998). Углеводы в отдельных клетках эпидермиса, мезофилла и пучкового влагалища листьев ячменя с измененной интенсивностью экспорта или фотосинтеза. Физиол растений. 118, 1525–1532. doi: 10.1104/pp.118.4.1525
Полный текст CrossRef | Google Scholar
Kusch, U., Greiner, S., Steininger, H., Meyer, A.D., Corbière-Divialle, H., Harms, K., et al. (2009). Анализ регуляции метаболизма фруктана в волосистых корнях цикория ( Cichorium intybus ). Новый Фитол. 184, 127–140. doi: 10.1111/j.1469-8137.2009.02924.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Lammens, W., Le Roy, K., Yuan, S., Vergauwen, R., Rabijns, A., Van Laere, A., et al. (2012). Кристаллическая структура 6-SST/6-SFT из Pachysandra terminalis , растительный биосинтезирующий фруктан фермент в комплексе с его акцепторным субстратом 6-кестозой. Завод J. 70, 205–219. doi: 10.1111/j.1365-313X.2011.04858.x
Завод J. 70, 205–219. doi: 10.1111/j.1365-313X.2011.04858.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ливингстон, Д. П. III. Хинча, Д.К., и Хейер, А.Г. (2009). Фруктан и его связь с устойчивостью растений к абиотическим стрессам. Сотовый. Мол. Жизнь наук. 66, 2007–2023 гг. doi: 10.1007/s00018-009-0002-x
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Монти А., Амадуччи М. Т. и Вентури Г. (2005). Реакция роста, газообмен листьев и накопление фруктанов у топинамбура ( Helianthus tuberosus L.) при воздействии различных водных режимов. евро. Дж. Агрон. 23, 136–145. doi: 10.1016/j.eja.2004.11.001
CrossRef Полный текст | Google Scholar
Мошен С., Радоник Л. М., Эренболгер Г. Ф., Фернандес П., Лиа В., Паньего Н. Б. и др. (2014). «Функциональная геномика и трансгенез применительно к селекции подсолнечника», в Подсолнухи , изд. JI Arribas (Nova: Science Publishers, Inc. ), 131–164.
), 131–164.
Google Scholar
Нелл, Дж. С. (2007). Генетические манипуляции с тканями, хранящими сахарозу, для производства альтернативных продуктов. кандидат наук диссертация, Стелленбосский университет, Стелленбос. Режим доступа: http://scholar.sun.ac.za/handle/10019.1/1359
Google Scholar
Парванова Д., Попова А., Захариева И., Ламбрев П., Константинова Т. , Танева С. и др. (2004). Низкотемпературная толерантность растений табака, трансформированных для накопления пролина, фруктанов или бетаина глицина. Свидетельство переменной флуоресценции хлорофилла. Фотосинтетика 42, 179–185. doi: 10.1023/B:PHOT.0000040588.31318.0f
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Pilon-Smits, E.A.H., Ebskamp, MJM, Paul, MJ, Jeuken, MJW, Weisbeek, PJ, and Smeekens, SCM (1995). Улучшение характеристик трансгенного табака, накапливающего фруктаны, в условиях засушливого стресса. Завод физиол. 107, 125–130. doi: 10.1104/pp.107.1.125
doi: 10.1104/pp.107.1.125
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Поллок С., Фаррар Дж., Томос Д., Галлахер Дж., Лу С. и Королева О. (2003). Баланс спроса и предложения: пространственная регуляция углеродного обмена в листьях трав и злаков. Дж. Экспл. Бот. 54, 489–494. doi: 10.1093/jxb/erg037
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Садрас, В. О., Коннор, Д. Дж., и Уитфилд, Д. М. (1993). Урожайность, компоненты урожайности и взаимосвязь между источником и стоком у подсолнечника, испытывающего дефицит воды. Полевые культуры Res. 31, 27–39. doi: 10.1016/0378-4290(93)
-R
CrossRef Full Text | Google Scholar
Садрас, В. О., и Холл, А. Дж. (1989). Характер водообеспеченности посевов подсолнечника в полузасушливой Центральной Аргентине. Основанная на моделировании оценка их взаимодействия со стратегиями возделывания культур и признаками сортов. С/х. Сист. 31, 221–238. doi: 10.1016/0308-521X(89)
doi: 10.1016/0308-521X(89)
-X
CrossRef Full Text | Google Scholar
Seiler, GJ (2007). Потенциал диких видов подсолнечника для промышленного использования. Гелия 30, 175–198. doi: 10.2298/HEL0746175S
Полный текст CrossRef | Google Scholar
Sims, D.A., Cheng, W., Luo, Y., и Seeman, JR (1999). Фотосинтетическая акклиматизация к повышенному содержанию CO2 в кроне подсолнуха. Дж. Экспл. Бот. 50, 645–653. doi: 10.1093/jxb/50.334.645
CrossRef Полный текст | Google Scholar
Tognetti, JA, Pontis, HG, and Martinez-Noël, GMA (2013). Передача сигналов сахарозой в растениях: мир, который еще предстоит изучить. Сигнал завода. Поведение 8, е23316. doi: 10.4161/psb.23316
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Валлору, Р., и Ван ден Энде, В. (2008). Фруктаны растений в условиях стресса: новые концепции и перспективы на будущее. Дж. Экспл. Бот. 59, 2905–2916. doi: 10.1093/jxb/ern164
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ван ден Энде, В.
Администрация муниципального образования Наурузовский сельсовет Пономаревского района Оренбургской области
При полном или частичном использовании
информации гиперссылка на nauruzovo.ru обязательна