Содержание
Жало осы — строение и интересные факты его применения
Содержание
Жало есть только у самок ос. Оно выполняет роль оружия защиты и нападения. С его помощью оса парализует жертву и уносит в гнездо для пропитания ее подрастающего потомства.
Некоторым людям вопрос о наличии жала у осы может показаться очевидным. Тем не менее, это не совсем так.
На самом деле жало осы есть у каждой самки, но самцы не могут похвастаться таким уникальным оружием. Уверенность людей в наличии жала у всех ос обусловлена тем, что большинство ос, относящихся к распространенному бумажному виду, являются самками. Именно поэтому жало есть в арсенале почти у всех ос, которых можно встретить на участке, балконе квартиры, в доме или просто на улице.
Функциональность жала
Жало для ос представляет собой основное оружие защиты от врагов и средство нападения на своих жертв. Хотя взрослые представители ос являются самыми настоящими вегетарианцами. Они предпочитают питаться нектаром, пыльцой цветущих растений, иногда высасывают кровь из своих жертв. Убитые насекомые нужны им для кормления личинок, только личинки способны питааться твердой пищей. Для добычи этой пищи и нужно жало.
Убитые насекомые нужны им для кормления личинок, только личинки способны питааться твердой пищей. Для добычи этой пищи и нужно жало.
Достаточно интересным фактом является то, что в период охоты осы предпочитают не слишком затрачивать имеющийся у них яд. С целью его экономии они убивают жертв своими мощными челюстями. перегрызая их тела. Даже самые плотные хитиновые покровы насекомого, которого оса решила убить, не станут серьезной проблемой.
Жало является универсальным оружием всех ос, и тех, что живут колониями, и ос-одиночек. Последние практически всегда используют его для того, чтобы обеспечивать свое потомство необходимыми запасами пищи. Все парализованные ядом пауки в последствии складываются в гнезда, где появившиеся на свет личинки без помощи матки сами справятся с пропитанием, ведь у них для этого будут достаточные запасы. Практически у всех ос жало характеризуется одинаковым строением. Разница может лишь заключаться в особенностях его использования и в составе яда.
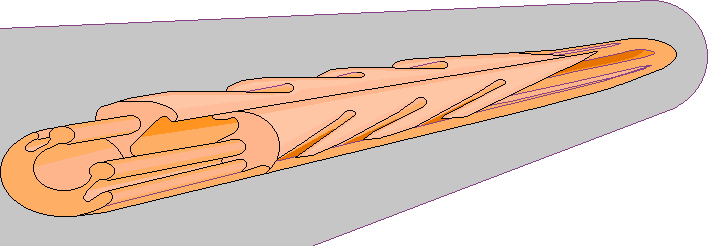
Строение жала осы
- ядовитая железа;
- заостренный хоботок;
- внутренний проток.
Стоит акцентировать внимание на третьем пункте. Специальный внутренний желобок жала предназначен для того, чтобы через него яд протекал и затем попадал в тело укушенной жертвы. Еще одной особенностью жала является его достаточно внушительная длина.
По своему строению жало осы отличается от жала пчелы Несмотря на свою визуальную тонкость, данное орудие является весьма прочным, т.к. по большей части состоит из хитина. По своим внешним параметрам жало вполне может напоминать самую обычную иглу. Формируется оно непосредственно из яйцеклетки. Этим как раз и объясняется факт того, что жало присутствует только у самок. У самцов попросту нет физиологических возможностей для его появления.
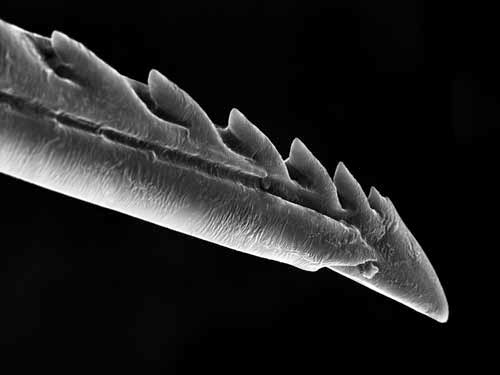
Жало под микроскопом
Как известно, главное орудие всех ос женского пола располагается в задней части брюшка. Если насекомое находится в спокойном состоянии и не чувствует приближающейся опасности, то жало может втягиваться. Правда, такой особенностью обладают не все виды. После совершения укуса жало выводится из тела жертвы благодаря напряжению группы особых мускул. Под микроскопом видны очень гладкие стенки жала. Само жало кажется прозрачным, хотя без микроскопа мы видим жало темно-коричневым или даже черным.
Правда, такой особенностью обладают не все виды. После совершения укуса жало выводится из тела жертвы благодаря напряжению группы особых мускул. Под микроскопом видны очень гладкие стенки жала. Само жало кажется прозрачным, хотя без микроскопа мы видим жало темно-коричневым или даже черным.
Наверняка многие люди задумывались о том, чем же все-таки отличается жало пчелы и осы. На самом деле такие различия существуют. У осы жало более гладкое. У пчелы же на жале имеется несколько зазубрин. Именно из-за них жало пчелы прочно застревает в теле жертвы. Вытащить его полностью пчела уже не может, что в конечном итоге приводит к ее гибели. Осы в данном плане более живучие. Они могут несколько раз использовать свое жало, оставаясь при этом полностью здоровыми.
Особенности применения жала
Чисто по внешним признакам достаточно сложно отличить между собой самку и самца осы, из-за чего сложно предугадать, какое насекомое способно ужалить человека. Лучше уделить внимание функциональности представленного органа. Перед атакой жало осы выдвигается из брюшка. Сразу же после атаки оса может нанести еще один удар, а только затем спрячет свое оружие в своеобразную «кобуру». Чем мягче покровы тела жертвы, тем легче осе будет проколоть их. Из-за твердости поверхности тела жуков осы предпочитают не охотиться на них. Большинство жуков надежно защищены крепкими надкрыльями, которые даже хитиновое жало осы не способно проткнуть.
Перед атакой жало осы выдвигается из брюшка. Сразу же после атаки оса может нанести еще один удар, а только затем спрячет свое оружие в своеобразную «кобуру». Чем мягче покровы тела жертвы, тем легче осе будет проколоть их. Из-за твердости поверхности тела жуков осы предпочитают не охотиться на них. Большинство жуков надежно защищены крепкими надкрыльями, которые даже хитиновое жало осы не способно проткнуть.
После укуса оса вводит в тело жертвы яд и вынимает жало. В зависимости от конкретной ситуации (это была защита или нападение) насекомое принимает решение, улететь подальше или снова совершить атаку. За одно нападение оса способна ужалить человека примерно 4-5 раз. Один укус приводит к тому, что в тело попадает около 0,3-0,4 мг ядовитого вещества. Самые крупные виды ос вводят до 0,7 мг яда. Единственным положительным моментом для человека при укусе является то, что осы сами вынимают свое жало. Поэтому практически всегда отсутствует необходимость извлекать его из тела.
Жало пчелы: где находится, как выглядит
Содержимое
- 1 Как выглядит жало пчелы
- 2 Где находится жало у пчелы
- 3 Оставляет ли пчела жало при укусе
- 4 Как жалит пчела
- 4.
 1 Как выглядит жало пчелы после укуса
1 Как выглядит жало пчелы после укуса - 4.2 Как вытащить жало после укуса
- 4.
- 5 Заключение
- 6 Отзывы
 1 Как выглядит жало пчелы после укуса
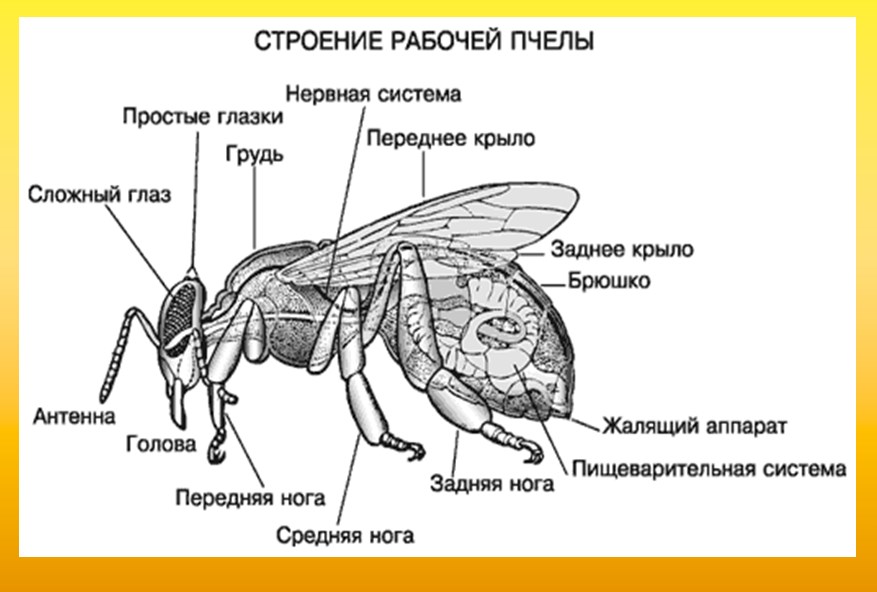
1 Как выглядит жало пчелы после укусаЖало пчелы – это орган, необходимый для защиты насекомым улья, используется только в случае возникновения опасности. Рассмотреть строение пчелиного жала в деталях можно при сильном увеличении под микроскопом. Расположено оно на кончике брюшка.
Как выглядит жало пчелы
Жалящий орган имеет сложное строение. Рассмотреть жало пчелы в деталях можно только при сильном увеличении под микроскопом: оно выглядит как острая длинная игла, утончающаяся от основания к кончику. По бокам хорошо заметны зазубрины, острыми концами направленные в сторону основания. У рабочих пчел их всего 10 штук, а у матки — 4. По сути, жало — это изменивший свое назначение яйцеклад. У трутней его нет совсем.
Оно состоит из нескольких элементов:
- трех хитиновых частей с пластинками;
- средняя часть — салазки, расширенная спереди и суженная сзади;
- стилет — состоит из двух ланцетов-игл, находится во впадине салазок снизу: при укусе стилет разламывается и освобождает иглы.

У каждой части органа свое предназначение. Стилетами насекомое пробивает кожу. Внутри салазок, в утолщенной части, расположена ядовитая железа, которая, в свою очередь, состоит из нитевидной доли и резервуара. В пузыре накапливается ядовитая жидкость. Рядом расположены железы, вырабатывающие смазку для стилета.
На фото под микроскопом можно видеть жало пчелы и извлеченное из тела укушенного человека инородное тело – ланцет:
Где находится жало у пчелы
Тельце насекомого разделено петиолью — талией — на грудку и брюшко. Верхнюю и нижнюю части в единый организм соединяет метасома — тонкий стебелек, через который проходят нервные окончания. Именно на кончике брюшка у пчелы есть жало. Его острие хорошо видно даже без сильного увеличения. Когда пчела спокойна, орган визуально незаметен.
Оставляет ли пчела жало при укусе
Орган после укуса застревает в ранке животного или человека. Проколов кожные покровы, стилет погружается в мягкий слой.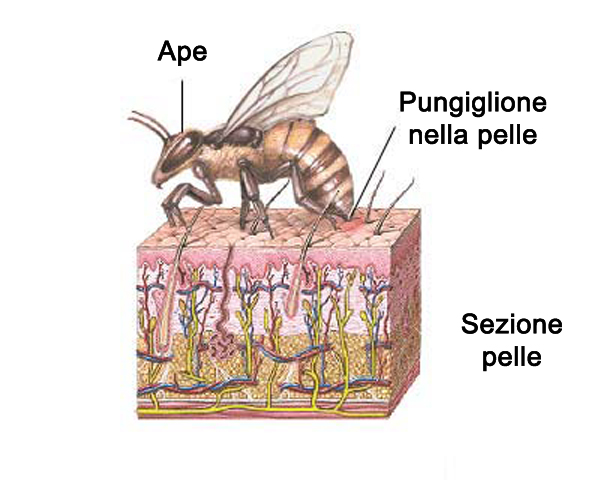 Инстинктивно пчела пытается вырвать стилеты из ранки, но зазубрины увязают в тканях. Жало отрывается от брюшка с частью внутренних органов. На тельце насекомого образуется рана, после чего оно погибает. Пчела выживает после схватки с осами и жуками. В хитиновых покровах зазубрины стилетов не застревают.
Инстинктивно пчела пытается вырвать стилеты из ранки, но зазубрины увязают в тканях. Жало отрывается от брюшка с частью внутренних органов. На тельце насекомого образуется рана, после чего оно погибает. Пчела выживает после схватки с осами и жуками. В хитиновых покровах зазубрины стилетов не застревают.
Как жалит пчела
В спокойном состоянии, когда насекомому ничего не угрожает, орган спрятан в специальном аппарате (мешке) на конце брюшка. Во время укуса жало из чехла выталкивается наружу. Мускулатура приводит в действие пластинки, вызывая скольжение стилетов по салазкам.
Подготавливаясь к атаке, пчела опускает жало вниз. Брюшко сильно прогибается вперед, а футляр приподнимается. В этот момент жалящий орган частично уже обнажен. Во время удара стилеты движутся резко вперед, затем мускулатура брюшка вбирает их обратно.
Пчела жалом пробивает поверхность кожи животного. После прокола в рану впрыскивается яд. Отравляющее вещество начинает стекать по салазкам.
Основной составляющей пчелиного яда является апитоксин: именно он вызывает жжение. Попавшее в организм вещество становится причиной болезненных ощущений. Кроме того, оно может стать поводом аллергических реакций. Ужаленные насекомые и мелкие животные (мыши) погибают от одной либо нескольких ядовитых инъекций. Единичный пчелиный укус большинству людей полезен. Многократно ужаленный человек получает слишком большую дозу яда. Смерть может наступить после накопления в организме 0,2 г апитоксина. Особенно опасны укусы в шею, глаза, губы.
Попавшее в организм вещество становится причиной болезненных ощущений. Кроме того, оно может стать поводом аллергических реакций. Ужаленные насекомые и мелкие животные (мыши) погибают от одной либо нескольких ядовитых инъекций. Единичный пчелиный укус большинству людей полезен. Многократно ужаленный человек получает слишком большую дозу яда. Смерть может наступить после накопления в организме 0,2 г апитоксина. Особенно опасны укусы в шею, глаза, губы.
Ядовитая жидкость имеет желтоватый оттенок. Попадая в кровь, яд быстро разносится по всему организму. Реакция на пчелиный укус у всех людей индивидуальна. В самых тяжелых случаях ядовитое вещество становится причиной возникновения у ужаленного человека:
- одышки;
- тошноты;
- головокружения;
- потери сознания;
- резких скачков артериального давления;
- отека дыхательных органов;
- покраснения части тканей, окружающих место прокола кожного покрова;
- болезненных ощущений;
- асфикции.

Как выглядит жало пчелы после укуса
После укуса оторванный орган начинает действовать автономно. Жало продолжает сокращаться, при этом выдавливая новые дозы яда в рану. Пульсируя, оно проникает еще глубже под кожу. Жало пчелы всей длиной погружается в ткани, и весь запас яда, который находится в мешочках в его основании, при сокращениях стекает по стилетам в образовавшийся канал, затем попадает в кровь. Область укуса очень быстро краснеет. Через некоторое время на поверхности остается видна только черная точка.
На фото видно жало пчелы, вырванное вместе с частью тельца насекомого, в коже человека. На поверхности заметна только верхняя часть органа: его остатки необходимо удалить как можно быстрее. Укушенное место выглядит опухшим, вокруг ранки быстро образуется отек. Посредине хорошо заметна черная точка.
Как вытащить жало после укуса
Опасность состоит в том, что поврежденное место быстро отекает и краснеет, и у укушенного человека может начаться аллергическая реакция.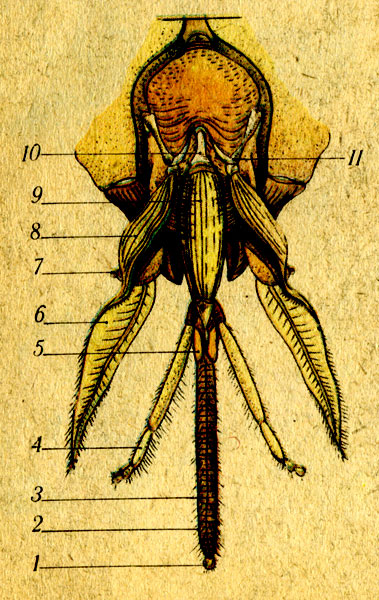 Жало, которое пчела оставляет в коже, продолжает поставлять в рану яд. Его необходимо удалить, однако нельзя это делать ногтями, иголкой, ножницами, а в стерильных условиях вытащить пинцетом, раскачивая из стороны в сторону. При удалении нужно убедиться, что вышел желтый мешочек с ядом на конце иглы. Если кусочек органа обломился и остался под кожей, придется обратиться за помощью к хирургу.
Жало, которое пчела оставляет в коже, продолжает поставлять в рану яд. Его необходимо удалить, однако нельзя это делать ногтями, иголкой, ножницами, а в стерильных условиях вытащить пинцетом, раскачивая из стороны в сторону. При удалении нужно убедиться, что вышел желтый мешочек с ядом на конце иглы. Если кусочек органа обломился и остался под кожей, придется обратиться за помощью к хирургу.
После удаления пчелиного жала место укуса нужно обработать антисептиком: спиртом, зеленкой, перекисью водорода и положить лед. Опытные пчеловоды при отсутствии аллергии советуют раствор меда для облегчения боли: разводят чайную ложку в стакане воды и выпивают. Для нейтрализации аллергена следует принять антигистаминный препарат.
Заключение
Жало пчелы необходимо насекомому, прежде всего, для защиты. Поэтому при столкновении с пчелой важно не провоцировать ее активными действиями (особенно, не махать руками), а постараться спокойно перейти в безопасное место. Укус малоприятен, но при отсутствии аллергии неопасен: важно полностью извлечь жало из-под кожи.
Отзывы
Светлана, 23 года, Москва
С опасностью, которую представляет жало пчел, мне довелось познакомиться в начале лета. Случилось так, что я получила укус прямо в верхнее веко. Жало быстро вытащили: кроме лёгкой припухлости на веке, меня ничего не беспокоило. Но следующий день заставил меня попереживать: утром я обнаружила, что полностью опухло и нижнее веко, глаз почти закрылся. Мы тут же вызвали «скорую» — оказалось, это был отек Квинке. От врачей узнала, что мне очень повезло и я легко отделалась, а ведь могла задохнуться. Поэтому когда вас укусит пчела, немедленно бегите к врачу, даже если симптомы незаметны.
Олег, 43 года, Пермь
Занимаюсь пчеловодством уже восьмой год. Некоторые считают, что если человека укусила пчела и несколько дней не проходит опухоль — значит, тело очищается. Не стоит верить этому: ведь пока что у организма просто нет привычки к яду. А если у человека имеется аллергия в опасной форме — могут возникнуть серьезные осложнения. Поэтому в случае укуса не стоит пренебрегать вызовом врача, это может спасти жизнь.
Структурные основы связывания STING с TBK1 и фосфорилирования с помощью TBK1
- Письмо
- Опубликовано:
- Conggang Zhang 1 na1 ,
- Guijun Shang 2 na1 ,
- Xiang Gui 1 ,
- Xuewu Zhang 2,3 ,
- Xiao-chen Bai 3,4 и
- …
- Чжицзянь Дж. Чен 1,5,6
Природа
том 567 , страницы 394–398 (2019)Цитировать эту статью
34 тыс.
обращений277 цитирований
120 Альтметрический
Сведения о показателях
 обращений
обращенийИспытуемые
- Криоэлектронная микроскопия
- Рецепторы распознавания образов
Abstract
Инвазия в цитоплазму млекопитающих микробной ДНК из инфекционных патогенов или собственной ДНК из ядра или митохондрий представляет собой сигнал опасности, который предупреждает иммунную систему хозяина 1 . Циклическая GMP-AMP-синтаза (cGAS) представляет собой сенсор цитоплазматической ДНК, который активирует путь интерферона I типа 2 . При связывании с ДНК cGAS активируется, чтобы катализировать синтез циклического GMP-AMP (cGAMP) из GTP и ATP 3 .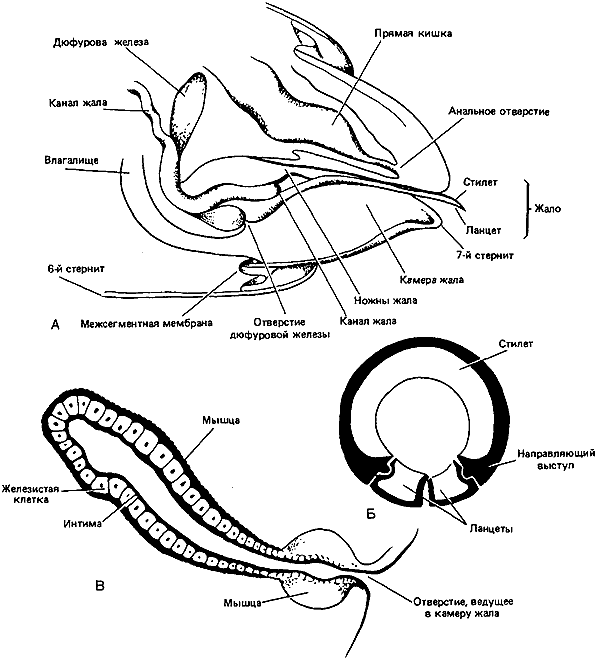 cGAMP функционирует как вторичный мессенджер, который связывается и активирует стимулятор генов интерферона (STING) 3,4,5,6,7,8,9 . Затем STING рекрутирует и активирует связывающую резервуар киназу 1 (TBK1), которая фосфорилирует STING и фактор транскрипции IRF3, чтобы индуцировать интерфероны типа I и другие цитокины 10,11 . Однако то, как STING, связанный с cGAMP, активирует TBK1 и IRF3, неясно. Здесь мы представляем структуру криоэлектронной микроскопии человеческого TBK1 в комплексе с цГАМФ-связанным полноразмерным куриным STING. Структура показывает, что C-концевой хвост STING принимает конформацию, подобную β-цепи, и вставляется в бороздку между киназным доменом одной субъединицы TBK1 и каркасом и доменом димеризации второй субъединицы в димере TBK1. В этом режиме связывания сайт фосфорилирования Ser366 в хвосте STING не может достичь активного сайта киназного домена связанного TBK1, что предполагает, что фосфорилирование STING с помощью TBK1 требует олигомеризации обоих белков.
cGAMP функционирует как вторичный мессенджер, который связывается и активирует стимулятор генов интерферона (STING) 3,4,5,6,7,8,9 . Затем STING рекрутирует и активирует связывающую резервуар киназу 1 (TBK1), которая фосфорилирует STING и фактор транскрипции IRF3, чтобы индуцировать интерфероны типа I и другие цитокины 10,11 . Однако то, как STING, связанный с cGAMP, активирует TBK1 и IRF3, неясно. Здесь мы представляем структуру криоэлектронной микроскопии человеческого TBK1 в комплексе с цГАМФ-связанным полноразмерным куриным STING. Структура показывает, что C-концевой хвост STING принимает конформацию, подобную β-цепи, и вставляется в бороздку между киназным доменом одной субъединицы TBK1 и каркасом и доменом димеризации второй субъединицы в димере TBK1. В этом режиме связывания сайт фосфорилирования Ser366 в хвосте STING не может достичь активного сайта киназного домена связанного TBK1, что предполагает, что фосфорилирование STING с помощью TBK1 требует олигомеризации обоих белков. Мутационный анализ подтверждает способ взаимодействия между TBK1 и STING и поддерживает модель, в которой олигомеризация STING и TBK1 высокого порядка, индуцированная cGAMP, приводит к фосфорилированию STING с помощью TBK1.
Мутационный анализ подтверждает способ взаимодействия между TBK1 и STING и поддерживает модель, в которой олигомеризация STING и TBK1 высокого порядка, индуцированная cGAMP, приводит к фосфорилированию STING с помощью TBK1.
Это предварительный просмотр содержимого подписки, доступ через ваше учреждение
Соответствующие статьи
Статьи открытого доступа со ссылками на эту статью.
Прерывание трафика STING после Гольджи активирует тоническую передачу сигналов интерферона.
- Синьтао Ту
- , Тин-Тин Чу
- … Нан Ян
Связь с природой
Открытый доступ
15 ноября 2022 г.Новая роль передачи сигналов STING при повреждении ЦНС: воспаление, аутофагия, некроптоз, ферроптоз и пироптоз
- Синьли Ху
- , Хаоцзе Чжан
- … Кайлян Чжоу
Журнал нейровоспаления
Открытый доступ
04 октября 2022 г.NAT10 регулирует пироптоз нейтрофилов при сепсисе посредством ацетилирования РНК ULK1 и активации пути STING.
- Хао Чжан
- , Чжаоюань Чен
- … Ванькун Чен
Биология коммуникаций
Открытый доступ
06 сентября 2022 г.

Варианты доступа
Подписаться на журнал
Получить полный доступ к журналу на 1 год
199,00 €
всего 3,90 € за выпуск
Подписаться
Расчет налога будет завершен во время оформления заказа.
Купить статью
Получите ограниченный по времени или полный доступ к статье на ReadCube.
32,00 $
Купить
Все цены указаны без учета стоимости.
Рис. 1: Структура комплекса куриного STING и человеческого TBK1. Рис. 2: Интерфейс связывания между человеческим TBK1 и С-концевым хвостом STING курицы. Рис. 3: Интерфейс между TBK1 и С-концевым хвостом STING необходим для связывания TBK1 и колокализации со STING в клетках. Рис. 4: Активация TBK1 и фосфорилирование STING зависят от олигомеризации STING.
Доступность данных
Крио-ЭМ-карта хвостового комплекса TBK1-STING депонирована в Банк данных электронной микроскопии (EMDB) под номером доступа EMD-0506. Атомные координаты комплекса депонированы в PDB под инвентарным номером 6NT9.
Ссылки
Li, T. & Chen, Z. J. Путь cGAS-cGAMP-STING связывает повреждение ДНК с воспалением, старением и раком. Дж. Эксп. Мед . 215 , 1287–1299 (2018).
Артикул
КАСGoogle ученый
Sun, L., Wu, J., Du, F., Chen, X. & Chen, Z. J. Циклическая GMP-AMP-синтаза представляет собой цитозольный ДНК-сенсор, который активирует путь интерферона I типа. Наука 339 , 786–791 (2013).
Артикул
ОБЪЯВЛЕНИЯ
КАСGoogle ученый
Ву, Дж. и др. Циклический GMP-AMP является эндогенным вторичным мессенджером в передаче сигналов врожденного иммунитета цитозольной ДНК. Наука 339 , 826–830 (2013).
Артикул
ОБЪЯВЛЕНИЯ
КАСGoogle ученый
Ishikawa, H. & Barber, G. N. STING — это адаптер эндоплазматического ретикулума, который способствует передаче сигналов врожденного иммунитета. Природа 455 , 674–678 (2008).
Артикул
ОБЪЯВЛЕНИЯ
КАСGoogle ученый
Сайто, Т. и др. Atg9a контролирует управляемую двухцепочечной ДНК динамическую транслокацию STING и врожденный иммунный ответ. Проц. Натл акад. науч. США 106 , 20842–20846 (2009 г.).
Артикул
ОБЪЯВЛЕНИЯ
КАСGoogle ученый
Джин, Л. и др. MPYS, новый мембранный тетраспаннер, связан с главным комплексом гистосовместимости класса II и опосредует передачу апоптотических сигналов. Мол. Клетка. Биол . 28 , 5014–5026 (2008 г.).
Артикул
КАСGoogle ученый
Burdette, D.L. et al. STING является датчиком прямого врожденного иммунитета циклического ди-ГМФ.
Природа 478 , 515–518 (2011).Артикул
ОБЪЯВЛЕНИЯ
КАСGoogle ученый
Zhang, X. et al. Циклический GMP-AMP, содержащий смешанные фосфодиэфирные связи, является эндогенным высокоаффинным лигандом для STING. Мол. Cell 51 , 226–235 (2013).
Артикул
КАСGoogle ученый
Кай, X., Чиу, Ю. Х. и Чен, З. Дж. Путь cGAS-cGAMP-STING восприятия цитозольной ДНК и передачи сигналов. Мол. Cell 54 , 289–296 (2014).
Артикул
КАСGoogle ученый
Лю, С. и др. Фосфорилирование адапторных белков врожденного иммунитета MAVS, STING и TRIF индуцирует активацию IRF3. Science 347 , aaa2630 (2015).
Артикул
Google ученый
Bai, X.C., Rajendra, E., Yang, G., Shi, Y. & Scheres, S.H. Выборка конформационного пространства каталитической субъединицы γ-секретазы человека. eLife 4 , e11182 (2015).
Артикул
Google ученый
Лараби, А. и др. Кристаллическая структура и механизм активации TANK-связывающей киназы 1. Cell Reports 3 , 734–746 (2013).
Артикул
КАСGoogle ученый
Ту, Д. и др. Структура и убиквитин-зависимая активация TANK-связывающей киназы 1. Cell Reports 3 , 747–758 (2013).
Артикул
КАСGoogle ученый
Чжао, Б. и др. Структурная основа согласованного рекрутирования и активации IRF-3 адапторными белками врожденного иммунитета. Проц. Натл акад. науч. США 113 , E3403–E3412 (2016 г.).
Артикул
КАСGoogle ученый
Лу, Д. и др. Структурное понимание эффекторного белка Tse3 Tse3 и комплекса Tse3-Tsi3 из Pseudomonas aeruginosa обнаруживают кальций-зависимый мембраносвязывающий механизм. Мол. Микробиол . 92 , 1092–1112 (2014).
Артикул
КАСGoogle ученый
Zheng, S.Q. et al. MotionCor2: анизотропная коррекция движения, вызванного лучом, для улучшения криоэлектронной микроскопии.
Нац. Методы 14 , 331–332 (2017).Артикул
КАСGoogle ученый
Чжан, К. Gctf: определение и коррекция CTF в реальном времени. Дж. Структура. Биол . 193 , 1–12 (2016).
Артикул
ОБЪЯВЛЕНИЯ
КАСGoogle ученый
Scheres, S.H. RELION: реализация байесовского подхода к определению структуры крио-ЭМ. Дж. Структура. Биол . 180 , 519–530 (2012).
Артикул
КАСGoogle ученый
Эмсли П., Локамп Б., Скотт В. Г. и Коутан К. Особенности и развитие Coot. Acta Кристаллогр. D 66 , 486–501 (2010).
Артикул
КАСGoogle ученый
Адамс, П. Д. и др. PHENIX: комплексная система на основе Python для решения макромолекулярной структуры.
Acta Кристаллогр. D 66 , 213–221 (2010).Артикул
КАСGoogle ученый
Чен В.Б. и др. MolProbity: проверка структуры всех атомов для макромолекулярной кристаллографии. Acta Кристаллогр. Д 66 , 12–21 (2010).
Артикул
КАСGoogle ученый
Петтерсен, Э. Ф. и др. UCSF Chimera — система визуализации для поисковых исследований и анализа. Дж. Вычисл. Химия . 25 , 1605–1612 (2004).
Артикул
КАСGoogle ученый
Гуэ, П., Курсель, Э., Стюарт, Д. И. и Метоз, Ф. ESPript: анализ множественных выравниваний последовательностей в PostScript. Биоинформатика 15 , 305–308 (1999).
Артикул
КАСGoogle ученый

 «>
«>Чжун Б. и др. Адаптерный белок MITA связывает рецепторы, чувствительные к вирусам, с активацией фактора транскрипции IRF3. Иммунитет 29 , 538–550 (2008 г.).
Артикул
КАС
Google ученый
 Природа 478 , 515–518 (2011).
Природа 478 , 515–518 (2011). «>
«>Шан Г., Чжан К., Чен З. Дж., Бай Х.-к. & Zhang, X. Крио-ЭМ структуры STING раскрывают его механизм активации с помощью циклического GMP-AMP. Природа https://doi.org/10.1038/s41586-019-0998-5 (2019).
 «>
«>Шу, К. и др. Структурное понимание функций TBK1 во врожденном противомикробном иммунитете. Структура 21 , 1137–1148 (2013).
Артикул
КАС
Google ученый
 Нац. Методы 14 , 331–332 (2017).
Нац. Методы 14 , 331–332 (2017). Acta Кристаллогр. D 66 , 213–221 (2010).
Acta Кристаллогр. D 66 , 213–221 (2010).Загрузить ссылки
Благодарности
Мы благодарим H. Yu и R. Hibbs за обмен инструментами и реагентами, а также X. Tan за вклад в анализ некоторых мутантов STING. Данные крио-ЭМ были собраны в Центре криоэлектронной микроскопии Юго-Западного медицинского центра Техасского университета (UTSW), который финансируется Техасским институтом профилактики и исследования рака (CPRIT) в рамках премии RP170644. Мы благодарим Д. Никастро за доступ к объекту и сбор данных. Эта работа частично поддерживается Медицинским институтом Говарда Хьюза (Z.J.C.), грантами Национального института здравоохранения (GM088197 и R35GM130289 до XZ), гранты фонда Welch (I-1389 до ZJC; I-1702 до XZ; I-1944 до X.-c.B) и гранты CPRIT (RP150498 до ZJC; RP160082 до X.-cB.) . X.-c.B. и Х.З. являются учеными Вирджинии Мерчисон Linthicum в области медицинских исследований в UTSW. Z.J.C. является исследователем Медицинского института Говарда Хьюза.
Yu и R. Hibbs за обмен инструментами и реагентами, а также X. Tan за вклад в анализ некоторых мутантов STING. Данные крио-ЭМ были собраны в Центре криоэлектронной микроскопии Юго-Западного медицинского центра Техасского университета (UTSW), который финансируется Техасским институтом профилактики и исследования рака (CPRIT) в рамках премии RP170644. Мы благодарим Д. Никастро за доступ к объекту и сбор данных. Эта работа частично поддерживается Медицинским институтом Говарда Хьюза (Z.J.C.), грантами Национального института здравоохранения (GM088197 и R35GM130289 до XZ), гранты фонда Welch (I-1389 до ZJC; I-1702 до XZ; I-1944 до X.-c.B) и гранты CPRIT (RP150498 до ZJC; RP160082 до X.-cB.) . X.-c.B. и Х.З. являются учеными Вирджинии Мерчисон Linthicum в области медицинских исследований в UTSW. Z.J.C. является исследователем Медицинского института Говарда Хьюза.
Информация для рецензентов
Nature благодарит Андреа Аблассер, Филипа Кранзуша и Осаму Нуреки за их вклад в рецензирование этой работы.
Информация об авторе
Примечания автора
Эти авторы внесли равный вклад: Conggang Zhang, Guijun Shang.
Авторы и организации
Кафедра молекулярной биологии Юго-западного медицинского центра Техасского университета, Даллас, Техас, США Техасский юго-западный медицинский центр, Даллас, Техас, США
Guijun Shang & Xuewu Zhang
Кафедра биофизики Юго-западного медицинского центра Техасского университета, Даллас, Техас, США Center, Dallas, TX, USA
Xiao-chen Bai
Центр исследований воспаления Техасского университета Southwestern Medical Center, Dallas, TX, USA
Zhijian J. Chen
Медицинский институт Говарда Хьюза, Юго-западный медицинский центр Техасского университета, Даллас, Техас, США
Zhijian J. Chen
Авторы
- Conggang Zhang
- в
PubMed Google Scholar - Guijun Shang
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Академия - Xiang Gui
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Xuewu Zhang
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Xiao-chen Bai
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Zhijian J. Chen
Посмотреть публикации автора
Вы также можете искать этого автора в
PubMed Google Scholar
 Chen
ChenВклады
Все авторы участвовали в разработке исследования, анализе данных и подготовке рукописи; Г.С. и К.З. подготовили образцы белка для крио-ЭМ; X.-c.B., Г.С. и X.Z. сбор данных, обработка изображений, определение структуры и анализ; Ч.З. и Х.Г. проводили функциональные анализы под наблюдением Z.J.C.
Авторы переписки
Переписка с
Сюэу Чжан, Сяо-чень Бай или Чжицзянь Дж. Чен.
Заявление об этике
Конкурирующие интересы
Авторы не заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Примечание издателя: Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Расширенные данные и таблицы
Расширенные данные Рис. 1 Очистка STING и TBK1 и характеристика их взаимодействия.

a , Связывание между очищенным человеческим STING и TBK1. Tsi3-меченый TBK1 был захвачен гранулами, конъюгированными с Tse3. Вытягивание STING с помощью TBK1 оценивали с помощью вестерн-блоттинга. ИБ, иммуноблоттинг; СТИНГ-ФЛ, полноразмерный СТИНГ; ЖАЛО-Δхвост, ЖАЛО(1–343). b , И человеческий, и куриный STING способны индуцировать фосфорилирование человеческого TBK1 в клетках. Клетки HeLa-C9 с неопределяемым эндогенным STING использовали для создания клеточных линий, стабильно экспрессирующих STING-Flag человека или STING-Flag курицы. Клетки стимулировали цГАМФ (1 мкМ) и анализировали на фосфорилирование TBK1 с помощью иммуноблоттинга. c , Гель-фильтрационная хроматография гибридного комплекса STING курицы и TBK1 человека. Данные являются репрезентативными для двух независимых экспериментов.
Расширенные данные Рисунок 2 Блок-схема обработки крио-ЭМ изображений для комплекса между куриным STING и человеческим TBK1.
a , репрезентативная микрофотография. b , Репрезентативные 2D-классы неповрежденного комплекса. c , Репрезентативные 2D-классы обработки изображений, ориентированной на TBK1. № > 3. d , f , Окончательные реконструкции неповрежденного комплекса STING–TBK1 ( d ) и уточнения, ориентированного на TBK1 ( f ), с цветами, основанными на локальном разрешении. e , g , Кривые FSC золотого стандарта окончательных 3D-реконструкций неповрежденного комплекса и уточнения, ориентированного на TBK1. h , Процедура обработки изображений.
b , Репрезентативные 2D-классы неповрежденного комплекса. c , Репрезентативные 2D-классы обработки изображений, ориентированной на TBK1. № > 3. d , f , Окончательные реконструкции неповрежденного комплекса STING–TBK1 ( d ) и уточнения, ориентированного на TBK1 ( f ), с цветами, основанными на локальном разрешении. e , g , Кривые FSC золотого стандарта окончательных 3D-реконструкций неповрежденного комплекса и уточнения, ориентированного на TBK1. h , Процедура обработки изображений.
Расширенные данные Рис. 3 Карты плотности образцов.
Карты плотности образцов показаны для С-концевого хвоста куриного STING и различных частей человеческого TBK1.
Расширенные данные Рис. 4 Структурное сравнение апо TBK1 (код PDB 4IM0) и TBK1, связанного со STING.
a , Общие конструкции TBK1. b , Увеличенное изображение областей TBK1, связанных с хвостом C-конца STING.
Расширенные данные Рис. 5 Сохранение последовательности TBK1 человека и курицы.
Остатки, идентичные в TBK1 обоих видов, окрашены в серый цвет. Неконсервативные остатки окрашены в красный цвет; неидентичные, но похожие остатки окрашены в розовый цвет.
Расширенные данные Рис. 6 Связывание и фосфорилирование STING с помощью TBK1 зависит от интерфейса между TBK1 и С-концевым хвостом STING и от олигомеризации STING.
a , Мутации TBK1-связывающих остатков в хвосте STING уменьшают cGAMP-индуцированное фосфорилирование как TBK1, так и STING. Постнуклеарный супернатант S1 от клеток HEK293T, которые экспрессировали либо STING дикого типа, либо мутанты, инкубировали с АТФ в присутствии или в отсутствие cGAMP и подвергали анализу иммуноблоттинга на pTBK1, pSTING и STING. b, c, Мутации TBK1-связывающих остатков в хвосте STING снижают cGAMP-индуцированное фосфорилирование STING (b), но не олигомеризацию STING (c). Те же образцы, что и в и , были разрешены с помощью нативных гелей и проанализированы с помощью иммуноблоттинга. d – f , Мутации на границе олигомеризации STING снижают олигомеризацию STING, индуцированную cGAMP, а также фосфорилирование TBK1 и STING. Мутанты основаны на сопроводительном документе о структурах полноразмерного STING 9.0012 12 . Анализы в d , e и f проводились таким же образом, как и в a , b и c соответственно. Показанные здесь данные являются репрезентативными как минимум для трех независимых биологических повторов.
d – f , Мутации на границе олигомеризации STING снижают олигомеризацию STING, индуцированную cGAMP, а также фосфорилирование TBK1 и STING. Мутанты основаны на сопроводительном документе о структурах полноразмерного STING 9.0012 12 . Анализы в d , e и f проводились таким же образом, как и в a , b и c соответственно. Показанные здесь данные являются репрезентативными как минимум для трех независимых биологических повторов.
Расширенные данные Рис. 7 Мультипликационная модель STING-опосредованной активации TBK1 и нижележащего сигнального пути.
Индуцированная cGAMP олигомеризация STING приводит к кластеризации TBK1 и транс-аутофосфорилированию. Активированный TBK1 фосфорилирует С-концевые хвосты STING, которые не связаны с бороздкой SDD-киназного домена в TBK1. Фосфорилированные хвосты STING рекрутируют IRF3, который фосфорилируется TBK1. Фосфорилированный IRF3 образует димер и перемещается в ядро, чтобы инициировать транскрипцию генов IFN.
Расширенные данные Рис. 8 Сбор данных и статистика модели.
a , Сбор данных и статистика уточнения модели. b , Кривые FSC между картами и моделью.
Дополнительная информация
Дополнительная информация
Этот файл содержит дополнительные результаты: Функциональная консервация между человеком и курицей STING.
Сводка отчетов
Дополнительные рисунки
Дополнительный рисунок 1: Исходные необрезанные изображения гелей или пятен.
Видео 1
Увеличенный вид интерфейса между хвостовой частью С-терминала STING и TBK1.
Права и разрешения
Перепечатка и разрешения
Об этой статье
Эта статья цитируется
Новая роль передачи сигналов STING при повреждении ЦНС: воспаление, аутофагия, некроптоз, ферроптоз и пироптоз
- Синьли Ху
- Хаоцзе Чжан
- Кайлян Чжоу
Журнал нейровоспаления (2022)
Геовременно-пространственный и причинно-следственный эпидемиологический обзор и обзор генотоксичности каннабиса, каннабидиола и каннабиноидов в США, выраженной в заболеваемости раком, 2003–2017 гг .
: часть 3 — пространственно-временной, многовариантный и причинно-следственный вывод и исследовательский анализ рака предстательной железы и яичников.- Альберт Стюарт Рис
- Гэри Кеннет Халс
Архив общественного здравоохранения (2022)
Роль TBK1 в патогенезе рака и противоопухолевом иммунитете
- Остин П. Рунде
- Райан Мак
- Дживан Чжан
Журнал экспериментальных и клинических исследований рака (2022)
Молекулярное клонирование, характеристика и функциональный анализ неохарактеризованного C11orf96 ген
- Хунзао Ян
- Цзе Чжу
- Гуанцин Лю
Ветеринарные исследования BMC (2022)
Крио-ЭМ структура активного бактериального комплекса филаментов TIR-STING
- Бенджамин Р. Морхауз
- Мэтью С. Дж. Йип
- Филип Дж. Кранзуш
Природа (2022)
- Бенджамин Р.
 : часть 3 — пространственно-временной, многовариантный и причинно-следственный вывод и исследовательский анализ рака предстательной железы и яичников.
: часть 3 — пространственно-временной, многовариантный и причинно-следственный вывод и исследовательский анализ рака предстательной железы и яичников. Морхауз
МорхаузКомментарии
Отправляя комментарий, вы соглашаетесь соблюдать наши Условия и Правила сообщества. Если вы обнаружите что-то оскорбительное или не соответствующее нашим условиям или правилам, отметьте это как неприемлемое.
RCSB PDB — 7SSM: кристаллическая структура укуса человека R232 в комплексе с соединением 11
Предыдущий ANTEXT
Содержание макромолекулы
- Веса Структура: 24,39 KDA и NBSP 9004
- .
- Количество депонированных остатков: 210 
- Уникальные белковые цепи: 1
Кристаллическая структура человеческого STING R232 в комплексе с соединением 11
WWPDB Validation & NBSP & NBSP 3D Report & NBSPFULL Report
Оценка качества структуры LIGAND. Черней, Э.С., Zhang, L., Lo, J., Huynh, T., Wei, D., Ahuja, Ahuja, Quesnelle, C., Schieven, G.L., Futran, A., Locke, G.A., L, L , Z.,  Monereau, L., Chaudhry, C., Blum, J., Li, S., Fereshteh, M., Li-Wang, B., Gangwar, S., Pan, C., Chong, C. , Zhu X., Posy, S.L., Sack, J.S., Zhang, P., Ruzanov, M., Harner, M., Akhtar,  Schroeder, G.M., Vite, G., Fink, B. (2022) J Med Chem  65 : 3518-3538 Идентификация агонистов пути стимулятора генов интерферона (STING) была областью интенсивных исследований из-за их способности усиливать врожденный иммунный ответ и иммуногенность опухоли в контексте иммуноонкологической терапии. Идентификация агонистов пути стимулятора генов интерферона (STING) была областью интенсивных исследований из-за их потенциала для усиления врожденного иммунного ответа и иммуногенности опухоли в контексте иммуноонкологической терапии. Первоначальные усилия по идентификации агонистов STING были сосредоточены на модификации 2′,3′-cGAMP ( 1 ) (эндогенный лиганд-активатор STING) и других близкородственных циклических динуклеотидов (CDN). Хотя эти усилия успешно идентифицировали новые CDN, которые вошли в клинику, их полезность в настоящее время ограничена пациентами с солидными опухолями, которым агонисты STING могут быть доставлены внутриопухолево. Здесь мы сообщаем об открытии уникального класса ненуклеотидных низкомолекулярных агонистов STING, которые демонстрируют противоопухолевую активность при внутриопухолевом введении на сингенной мышиной модели. Организационная принадлежность :  Bristol Myers Squibb Research and Development, P.O. Box 4000, Принстон, Нью-Джерси 08543, США. Макромолекулы Найдите похожие белки по: (по порогу идентичности) | Трехмерная структура Найти белки для &nbbQ86WV0107 (Homo sapiens) Explore Q86WV6  Go to UniProtKB:  Q86WV6 PHAROS:  Q86WV6 Small Molecules Загрузить CCD-файл с идеальными координатами  Experimental Data & Validation Unit Cell : Software Package: View Full Validation Report View more in-depth experimental data Entry History & Funding Information Депонирование Автор(ы):  Сак, Дж. С., Криттон, Д. А. (Full details and data files)  См. полную историю.
См. полную историю. Первоначальные попытки идентифицировать агонисты STING были сосредоточены на модификации 2′,3′-cGAMP (9).0187 1 ) (эндогенный лиганд активатора STING) и другие близкородственные циклические динуклеотиды (CDN) …
Первоначальные попытки идентифицировать агонисты STING были сосредоточены на модификации 2′,3′-cGAMP (9).0187 1 ) (эндогенный лиганд активатора STING) и другие близкородственные циклические динуклеотиды (CDN) …
ID объекта: 1
Молекула Цепи Длина последовательности 1 Организм Details Image Stimulator of interferon genes protein A 210 Homo sapiens Mutation(s) : 0 
Gene Names:  STING, LOC340061, hCG_1782396, STING1, ERIS , MITA, TMEM173 Ресурсы данных Общего фонда UniProt и NIH
Entity Groups
& NBSP кластеры последовательности 30% идентичность.  0795
0795 Expand
Ligands 1 Unique ID Chains Name / Formula / InChI Key 2D Diagram 3D-взаимодействия B7L (предмет исследования/LOI)
Запрос на B7L B [auth A] 2-({[(8R)-пиразоло[1,5-a]пиримидин-3-карбонил]амино}метил)-1-бензофуран-7-карбоновая кислота
C 17 H 12 N 4 O 4
BICDHWPHEXMVAT-UHFFFAOYSA-N Ligand Interaction Experimental Data
Длина (Å) Angle ( ˚ ) a = 110.  08
08 α = 90 b = 110.08 β = 90 c = 36.2 γ = 90 Software Name Purpose Aimless data scaling BUSTER refinement PDB_EXTRACT data extraction PHASER phasing Structure Validation
Ligand Structure Quality Assessment 
Deposition Data
Funding Organization Location Grant Number Not funded — Revision History