Содержание
ФАЗА «КУЩЕНИЕ – ВЫХОД В ТРУБКУ»
Фазы роста и развития ячменя
Кущение. Что происходит с культурой эту фазу?
Кущение – это появление новых побегов у зерновых за счет подземного ветвления стебля.
Зона кущения является:
- Важным источником меристаметических тканей для формирования боковых и придаточных корней;
- Механической основой для надземной части побега;
- Распределителем воды и минеральных солей, поступающих из корней, и органических соединений, поступающих из листьев;
- Запасающим резервуаром питательных веществ.
Отмирание узла кущения приводит к гибели.
Как происходит кущение?
Почка, лежащая у основания первого листа, увеличивается, отодвигает лист и формирует первый боковой побег. В дальнейшем в пазухах нижних листьев боковых побегов закладываются новые почки, которые могут давать побеги 2-го, 3-го и большего порядка. Одновременно с образованием боковых побегов из узла кущения формируется вторичная корневая система. Вторичная корневая система хорошо развивается при наличии почвенной влаги и при наличии доступной формы фосфора. В сухом верхнем слое вторичные стебли и корни не образуются. К концу кущения растения ячменя поглощают около 50% азота и фосфора и 75% калия от общего потребления.
Одновременно с образованием боковых побегов из узла кущения формируется вторичная корневая система. Вторичная корневая система хорошо развивается при наличии почвенной влаги и при наличии доступной формы фосфора. В сухом верхнем слое вторичные стебли и корни не образуются. К концу кущения растения ячменя поглощают около 50% азота и фосфора и 75% калия от общего потребления.
Что влияет на глубину залегания узла кущения?
- Свет.При недостатке света узел кущения залегает ближе к поверхности.
- Глубина заделки семян, тип почвы.
- Температура. При пониженной температуре воздуха узел кущения заглубляется.
- Сорт.
Во время кущения происходят закладка побегов, колосков и цветочков, а также обильный рост корней.
Кущение – самая УЯЗВИМАЯ СТАДИЯ развития ячменя.
ЧТО МОЖЕТ НАНЕСТИ ВРЕД КУЛЬТУРЕ В ЭТУ ФАЗУ?
В фазу кущения посевам ячменя кроме ранних листовых вредителей, таких как блошка и др. листоеды, большой вред наносят сорняки. Всходы многих сорняков появляются одновременно со всходами ячменя и являются конкурентами за влагу и питательные вещества.
листоеды, большой вред наносят сорняки. Всходы многих сорняков появляются одновременно со всходами ячменя и являются конкурентами за влагу и питательные вещества.
Что может сделать агроном для будущего урожая прямо сейчас?
Подобрать гербициды с учетом видового состава сорняков и степени засоренности.
Химический метод – один из наиболее эффективных. При выборе гербицида для конкретного поля следует учитывать его эффективность к преобладающим там сорнякам. На одно поле может присутствовать более 20 видов сорняков из разных биологических групп. Показателем уровня засоренности следует считать не количество растений сорняков, а их массу на единицу площади. В этом отношении все сорняки условно можно разделить на доминанты, субдоминанты и сопутствующие виды.
Обработку против сорняков нужно проводить до экранирования сорняков культурой, т.е. гербициды должны попасть на целевой объект – сорняки.
При преобладании в посевах таких сорняков как молочай лозный, вьюнок полевой, марь белая и др.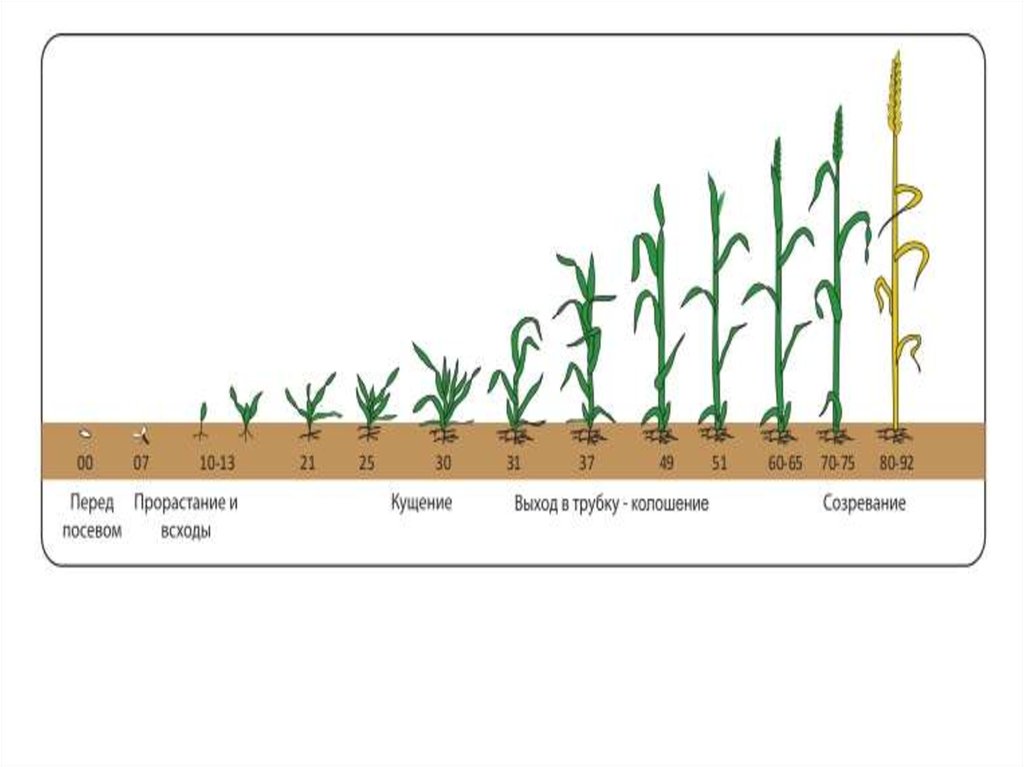 рекомендуется применять гормональные гербициды такие как Видмастер, Диален Супер, Линтур, которые содержат 2,4-Д и дикамбу. Действие таких препаратов основано на росторегулирующих функциях. Препарат проникает в сорное растение через листья и корни и распространяется системно по всему растению к точкам роста. В результате воздействия двух действующих веществ нарушается процесс нормального деления клеток, что приводит к деформации и скручиванию стеблей и листьев, а впоследствии к гибели растений.
рекомендуется применять гормональные гербициды такие как Видмастер, Диален Супер, Линтур, которые содержат 2,4-Д и дикамбу. Действие таких препаратов основано на росторегулирующих функциях. Препарат проникает в сорное растение через листья и корни и распространяется системно по всему растению к точкам роста. В результате воздействия двух действующих веществ нарушается процесс нормального деления клеток, что приводит к деформации и скручиванию стеблей и листьев, а впоследствии к гибели растений.
Препараты, содержащие в своем составе 2,4-Д и дикамбу применяют только в фазу кущения, до выхода в трубку.
Особое внимание необходимо уделять борьбе со злаковыми сорняками, такими как овсюг обыкновенный, щетинники и просовидные. Семена просовидных сорняков и щетинников легко отделяются от ячменя при очистке, тогда как очистить ячмень от овсюга тяжело, для этого необходимы специальные овсюжные триеры.
При смешанном типе засоренности рекомендуется применять баковые смеси противодвуольных и противозлаковых сорняков.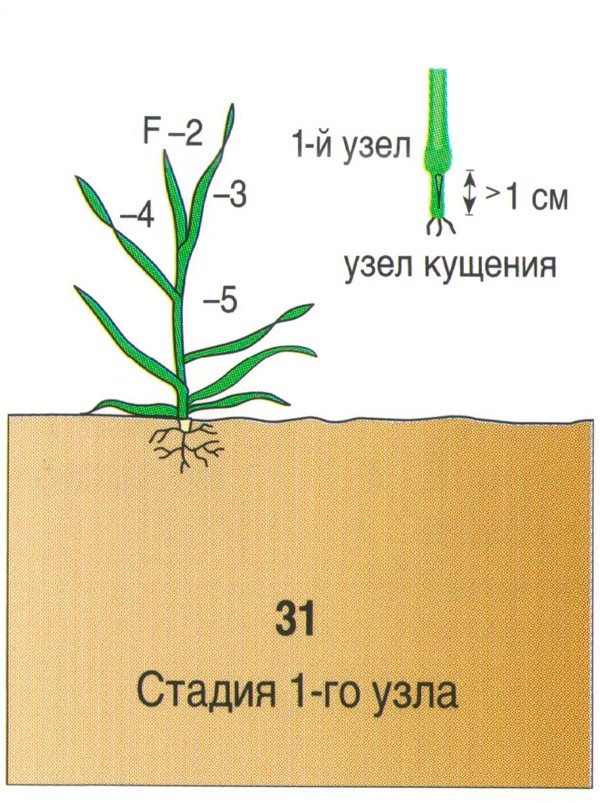 Использование баковых смесей дает фермерам возможность сократить расходы, сэкономить время и повысить биологическую и экономическую эффективность применения средств защиты растений (СЗР), удобрений. При смешивании баковых смесей необходимо соблюдать порядок смешивания препаратов, учитывающий их физико-химические свойства.
Использование баковых смесей дает фермерам возможность сократить расходы, сэкономить время и повысить биологическую и экономическую эффективность применения средств защиты растений (СЗР), удобрений. При смешивании баковых смесей необходимо соблюдать порядок смешивания препаратов, учитывающий их физико-химические свойства.
Выход в трубку. Что происходит с культурой в эту фазу?
Эта фаза активного роста ячменя.
Начало трубкования растений ячменя отмечается когда стеблевой узел прощупывается сквозь влагалище листа на высоте 3-5 см над поверхностью почвы. Нижние междоузлия раздвигаются и стебель начинает удлиняться. Рост стебля начинается с удлинения нижнего междоузлия. Рост междоузлий заканчивается к концу цветения – началу налива.
Фаза выхода в трубку длится в среднем 15-25 дней.
В этой фазе ячмень очень чувствителен к недостатку влаги, питательных веществ и других факторов жизни. Благоприятная температура в этот период +12-18˚С. К началу фазы трубкования ячмень поглощает до 70% калия, около 45% фосфора и значительную часть азота от общего количества используемого в течение вегетационного периода.
В межфазный период выход в трубку – колошение увеличивается площадь листовой поверхности, растет соломина. В этот период формируются и растет колос.
ЧТО МОЖЕТ НАНЕСТИ ВРЕД КУЛЬТУРЕ В ЭТУ ФАЗУ?
В фазу выхода в трубку посевам пивоваренного ячменя могут нанести вред болезни и вредители. В результате вреда таких вредителей как пшеничный трипс, злаковая тля колос деформируется, образуется белоколосость.
Пшеничный трипс Желтая ржавчина Ячменная муха Полосатая пятнистость
Засуха в этот период приводит к редукции, т.е. «сбросу» числа побегов и количества зерен в колосе.
Что может сделать агроном для будущего урожая прямо сейчас?
- Прекратить все гербицидные обработки. В эту фазу закладывается колос и применение повышенных норм гербицидов группы 2,4-Д в этот период может привести к частичной стерильности колоса.

- Провести инсектицидную обработку против вредителей.
- Провести профилактическую фунгицидную обработку против листо-стебельных болезней.
- Провести листовую подкормку удобрениями, содержащими

Кущение | это… Что такое Кущение?
Ячмень обыкновенный (лат. Hordéum vulgáre) в фазу кущения
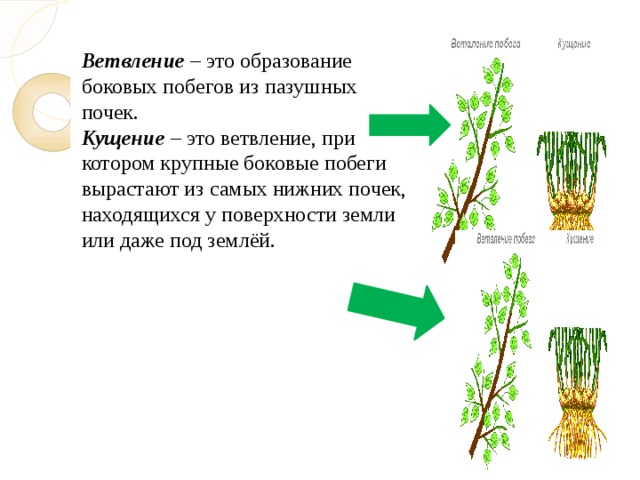
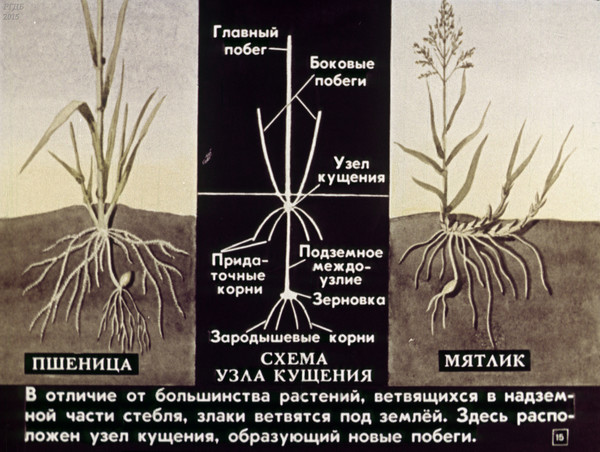
Куще́ние — образование надземных побегов из узла, расположенного у основания главного побега у растений семейства Злаки и некоторых других растений[1]; обильное появление у Злаков побочных стеблей (боковых побегов)[2], благодаря благоприятным условиям роста[3]; одна из форм ветвления, приводящая к образованию куста[4].
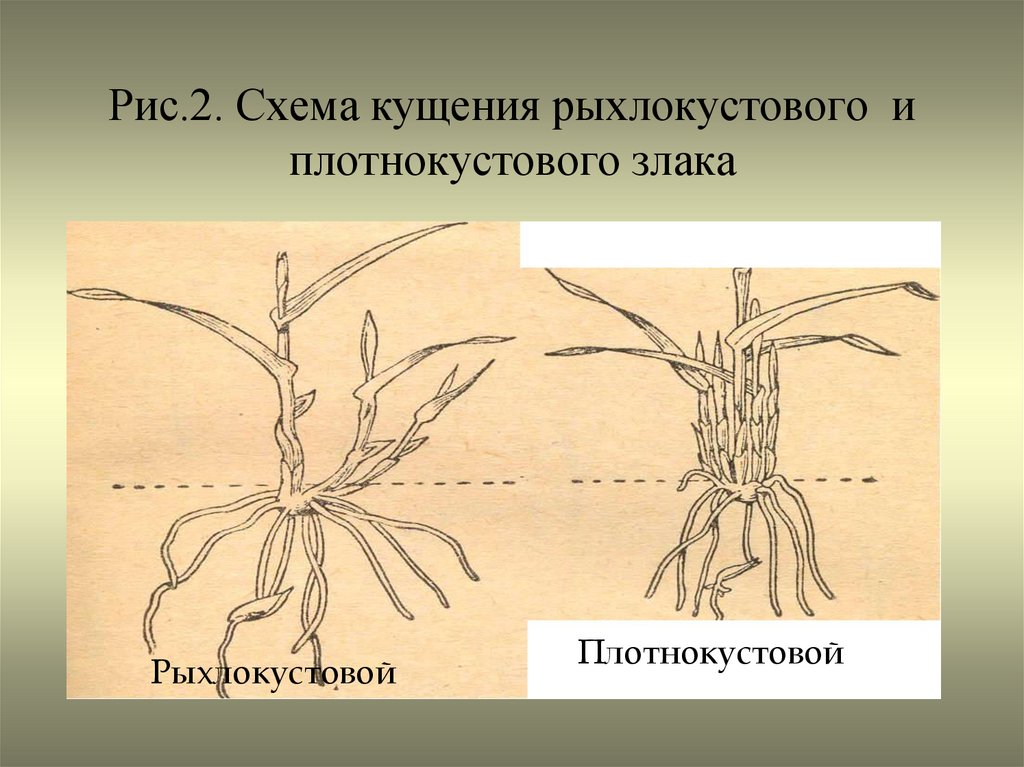
Узел кущения — это ряд сближенных коротких междоузлий, из почек которых развиваются боковые побеги[1][5]; из почек, которые находятся в пазухах влагалищных листьев, образуются дочерние боковые побеги, которые либо остаются внутри влагалища (внутривлагалищные побеги), либо пробивают влагалище листа и выходят наружу (вневлагалищные побеги)[1]. В результате образуется рыхлый (при условии, что побеги растут под углом к главному) или плотный (при условии, что побеги растут вертикально) куст[1][6].
В результате образуется рыхлый (при условии, что побеги растут под углом к главному) или плотный (при условии, что побеги растут вертикально) куст[1][6].
Из-за развития из узла кущения множества побегов многолетние растения образуют дернину; расположенные горизонтально длинные побеги при подземном их расположении образуют корневища или отпрыски при наземном их расположении[1]. У однолетних растений процесс кущения происходит в ранний период жизни и прекращается с выходом их в трубку[1][4]; у многолетних растений процесс кущения продолжается до конца вегетации после перерыва в период цветения и плодоношения[1]. У культурных растений кущение зависит от сорта, условий произрастания и агротехнических приёмов[4]. Кущение хлебных Злаков является одной из фаз развития, которая способствует увеличению урожая[6].
Примечания
- ↑ 1 2 3 4 5 6 7 Большая советская энциклопедия: В 30 т. — М.: «Советская энциклопедия», 1969—1978.
- ↑ Энциклопедический словарь Ф. А. Брокгауза и И. А. Ефрона. — С.-Пб.: Брокгауз-Ефрон. 1890—1907.
- ↑ Малый энциклопедический словарь Брокгауза и Ефрона/ В современной орфографии. — Петербург: Издательское общество «Ф. А. Брокгауз — И. А. Ефрон», 1907—1909.
- ↑ 1 2 3 Биологический энциклопедический словарь / Гл. ред. М. С. Гиляров; Редкол.: А. А. Баев, Г. Г. Винберг, Г. А. Заварзин и др. — М.: Сов. энциклопедия, 1986. — С. 307. — 100 000 экз.
- ↑ Биология. Современная иллюстрированная энциклопедия. Гл. ред. А. П. Горкин. — М.: Росмэн, 2006.
- ↑ 1 2 Большой Энциклопедический словарь. — 2000.
 — М.: «Советская энциклопедия», 1969—1978.
— М.: «Советская энциклопедия», 1969—1978.Литература
- Большая советская энциклопедия: В 30 т. — М.: «Советская энциклопедия», 1969—1978.
- Энциклопедический словарь Ф. А. Брокгауза и И. А. Ефрона. — С.-Пб.: Брокгауз-Ефрон. 1890—1907.
- Малый энциклопедический словарь Брокгауза и Ефрона/ В современной орфографии. — Петербург: Издательское общество «Ф. А. Брокгауз — И. А. Ефрон», 1907—1909.
- Биологический энциклопедический словарь / Гл. ред. М. С. Гиляров; Редкол.: А. А. Баев, Г. Г. Винберг, Г. А. Заварзин и др. — М.: Сов. энциклопедия, 1986. — С. 831. — 100 000 экз.
- Биология. Современная иллюстрированная энциклопедия. Гл. ред. А. П. Горкин. — М.: Росмэн, 2006.
- Большой Энциклопедический словарь. — 2000.
- Bell, Adrian (1991). Plant Form. Oxford, UK: Oxford University Press. pp. 182. ISBN 0-19-854219-4.
 Ефрона. — С.-Пб.: Брокгауз-Ефрон. 1890—1907.
Ефрона. — С.-Пб.: Брокгауз-Ефрон. 1890—1907.Кущение | Информационная система для кормов
Культиваторы очень важны для понимания роста и отрастания травы. К сожалению, этот термин имеет много синонимов и иногда сбивает с толку. Кусты — это новые побеги травы, состоящие из последовательных сегментов, называемых фитомерами, которые состоят из точки роста (апикальной меристемы, которая может превратиться в головку семени), стебля, листьев, корневых узлов и скрытых почек; все они могут расти из почек ткани кроны, корневищ, столонов или надземных узлов (воздушные побеги).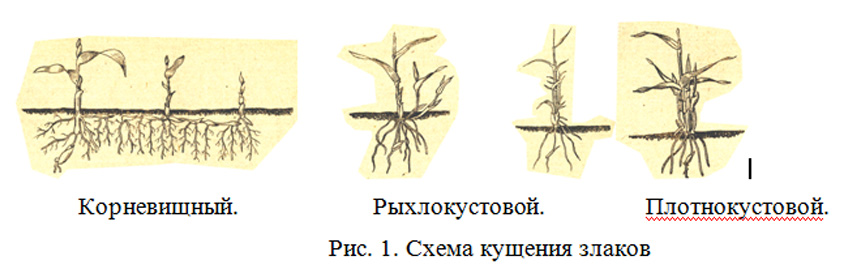
Кустик может зацвести, если его подвергнуть необходимым условиям выращивания, в противном случае он останется вегетативным. Процесс появления этих новых надземных побегов называется кущением. В отличие от появления корневищ и столонов, побеги развиваются вверх. Результатом является резкое увеличение количества новых побегов, возникающих непосредственно рядом с первоначальным побегом. Исходный побег иногда называют «материнским растением», а новые побеги — «дочерними растениями». Чепмен (1998) определил румпель как дополнительный стебель, возникающий у основания основного стебля или рядом с ним, или один из его более ранних ответвлений».
Одиночное зерно пшеницы может дать дополнительные побеги (кусты) из придаточных почек в зоне кроны. Кукуруза (кукуруза), однолетняя трава теплого сезона, производит присоски из базальных узлов. У многолетних трав некоторые виды образуют побеги из адвентивных почек, как и у кукурузы. Если начальный отросток остается внутри влагалища, окутывающего узел, ветвление классифицируется как интравагинальное. Примерами являются ежа, овсяница и другие дерновинные злаки. Если внутривлагалищный побег возникает выше на стебле (второй или третий узел от кроны), новый побег считается надземной ветвью, как у райграса однолетнего. Больше этой информации можно найти в разделе летних ежегодников.
Примерами являются ежа, овсяница и другие дерновинные злаки. Если внутривлагалищный побег возникает выше на стебле (второй или третий узел от кроны), новый побег считается надземной ветвью, как у райграса однолетнего. Больше этой информации можно найти в разделе летних ежегодников.
Если начальный побег растет латерально таким образом, что разрывает покрывающую его оболочку, то характер ветвления вневагинальный. Последняя классификация включает корневищные и столоносные злаки. Если корневища короткие, как у овса-грамы, трава не будет инвазивной для других видов.
Кущение может также относиться к стадии роста, когда появляются побеги. У злаковых культур ранневесеннее кущение называется «стеливанием». Каждый новый побег содержит центральную точку роста (зачаток побега), которая в конечном итоге превращается в сочлененный стебель, характеризующийся отчетливыми узлами и междоузлиями, как на бамбуковом шесте. Членистый стебель называется стеблем. Каждый узел стебля несет лист (лопасть и ножны). Самый верхний сегмент стебля, поддерживающий головку семени, называется цветоносом.
Самый верхний сегмент стебля, поддерживающий головку семени, называется цветоносом.
При кормовых посадках отдельные побеги в конечном итоге отмирают и должны быть заменены новыми побегами для поддержания желаемой густоты растений. Кормовые многолетние травы являются многолетними не потому, что отдельные побеги выживают бесконечно, а потому, что растительное сообщество динамично, и отмирающие члены постоянно заменяются новыми побегами. Срок жизни отдельного побега обычно не превышает одного года, а часто и меньше. Побеги, образующиеся осенью, важны для зимнего выживания насаждения и весеннего отрастания, но летом могут погибнуть. Кусты, сформированные весной, могут иметь важное значение для выживания летом. Побеги, дающие начало цветению весной, обычно отмирают до конца лета.
Молодой куст зависит от родительского побега в плане фотоассимиляции, пока у него не разовьется несколько листьев и адекватная корневая система. Хотя может показаться, что зрелый побег функционирует как независимая единица, очевидно, существует некоторая связь между побегами, связанными общей сосудистой системой. Таким образом, травянистое растение представляется скорее высокоорганизованной системой, чем набором конкурирующих побегов.
Таким образом, травянистое растение представляется скорее высокоорганизованной системой, чем набором конкурирующих побегов.
Последствия для управления
Кущение обеспечивает лучшее укоренение дерновинных злаков и боковое распространение (через корневища/столоны) дернообразующих злаков. Кусты дают много дополнительных корней. Поэтому весной следует отложить выпас скота до тех пор, пока они не смогут переносить ограниченную дефолиацию.
Практическое применение
1) Виды, которые имеют экстенсивное кущение, могут быть высажены с меньшей нормой высева.
2) При соблюдении определенных мер предосторожности ранний выпас не представляет серьезной угрозы для животновода. Например, фермеры, выращивающие пшеницу на Среднем Западе и Юге, обычно выпасают ранний рост пшеницы в фазе кущения. Однако, когда побеги начинают формировать центральный стебель (стадия раннего отсоединения или перехода), они перестают питаться из-за возможного разрушения точки роста, содержащей рудиментарную семенную головку.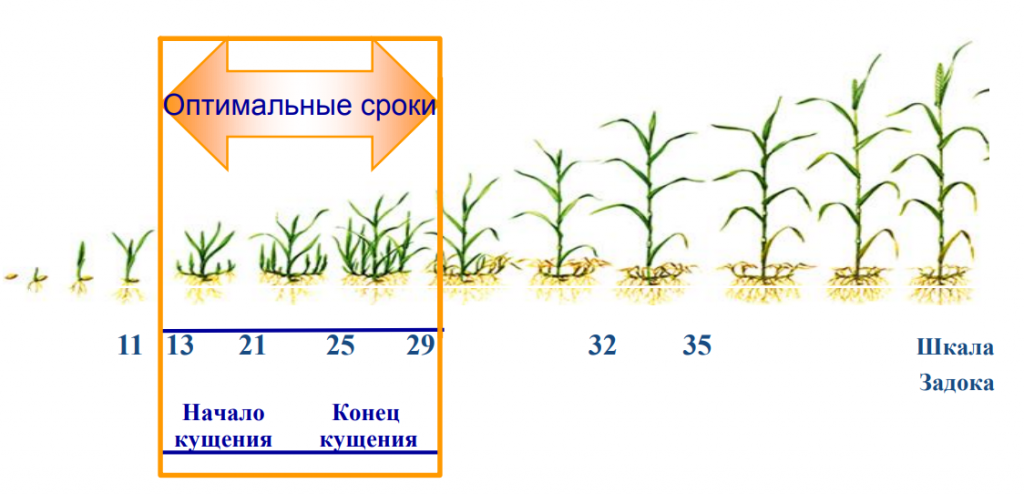 Точно так же в западном Орегоне овец обычно пасут на посевных полях травы, но убирают перед удлинением стебля. Но на этом этапе роста есть дополнительная угроза. Домашний скот может удалять листья с побега таким образом, чтобы сохранить точку роста, но может привести к удалению одной или нескольких листовых пластинок ниже воротниковой зоны. Когда это происходит, восстановительный рост состоит из цветущего стебля с неповрежденной головкой семени, но с сильно уменьшенной площадью листовой пластинки. Листья являются местом фотосинтеза, поэтому домашний скот не должен пастись так долго, чтобы растения оголились.
Точно так же в западном Орегоне овец обычно пасут на посевных полях травы, но убирают перед удлинением стебля. Но на этом этапе роста есть дополнительная угроза. Домашний скот может удалять листья с побега таким образом, чтобы сохранить точку роста, но может привести к удалению одной или нескольких листовых пластинок ниже воротниковой зоны. Когда это происходит, восстановительный рост состоит из цветущего стебля с неповрежденной головкой семени, но с сильно уменьшенной площадью листовой пластинки. Листья являются местом фотосинтеза, поэтому домашний скот не должен пастись так долго, чтобы растения оголились.
Вышеупомянутая реакция на ранний выпас обычно наблюдается при ротационном выпасе, когда несколько загонов выпасаются последовательно. Первый загон часто слишком долго выпасают, что приводит к восстановительному росту, состоящему в основном из скелетированного цветущего стебля (голый стебель с семенной головкой, но без листовых пластинок). Пастбище следует обрезать, чтобы уничтожить семенные головки и удалить апикальное доминирование, что позволит развиться базальным почкам, по существу направляя энергию растений на отрастание.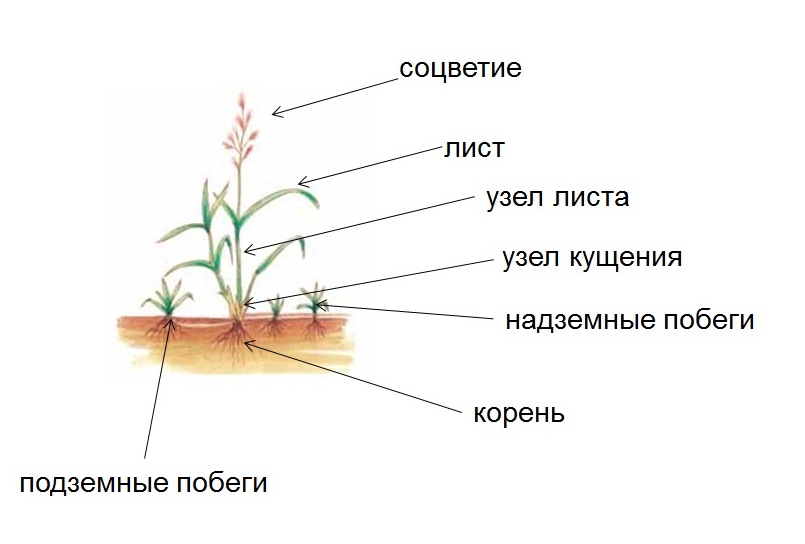
Если скоту будет позволено слишком интенсивно пастись во 2-м загоне, надземный механизм отрастания может быть полностью разрушен. Если этот эффект совпадет с коротким периодом, когда адвентивные почки в кроне плохо развиты и неспособны дать новые побеги, сорняки и бобовые будут процветать.
Продолжая описанную выше последовательность выпаса, загон № 3 будет выпасаться во время большой фазы роста (стадия позднего или раннего выхода в трубку). Трава близка к пику качества и количества. Если уровень посадки не увеличить, чтобы максимизировать прибыль от этого ресурса, многие побеги травы будут давать семенные головки, что сделает корм неприятным на вкус. Такие пастбища следует подстригать при появлении всходов. Это вызывает раннее отрастание, давая возможность второму циклу кущения создать новую корневую систему до наступления летней жары и засухи.
Контроль кущения риса
- Опубликовано:
- Xueyong Li 1 na1 ,
- Qian Qian 2 na1 ,
- Zhiming Fu 1 ,
- Yonghong Wang 1 ,
- Guosheng Xiong 1 ,
- Dali Zeng 2 ,
- Сяоцюнь Ван 1 ,
- Синьфан Лю 1 ,
- Sheng Teng 2 ,
- Fujimoto Hiroshi 2 ,
- Ming Yuan 3 ,
- Da Luo 4 ,
- Bin Han 5 na1 &
- …
- Цзяян Ли 1
Природа
том 422 , страницы 618–621 (2003 г. )Процитировать эту статью
)Процитировать эту статью
13 тыс. обращений
777 цитирований
7 Альтметрический
Сведения о показателях
Abstract
Кущение риса ( Oryza sativa L.) является важным агрономическим признаком для производства зерна, а также модельной системой для изучения ветвления однодольных растений. Побег риса — специализированная зерновая ветвь, образующаяся на неудлиненном базальном междоузлии и растущая независимо от материнского стебля (стебля) за счет собственных придаточных корней 1 . Кущение риса происходит в два этапа: образование пазушной почки в каждой пазухе листа и последующее ее отрастание 2 . Хотя морфология и гистология 2,3 и некоторых мутантов кущения риса 4 хорошо описаны, молекулярный механизм кущения риса еще предстоит выяснить. Здесь мы сообщаем о выделении и характеристике MONOCULM 1 ( MOC1 ), гена, важного для контроля кущения риса. Мутантные растения moc1 имеют только главный стебель без побегов из-за дефекта формирования почек побегов. MOC1 кодирует предполагаемый ядерный белок семейства GRAS, который экспрессируется в основном в пазушных зачатках и функционирует, чтобы инициировать пазушные зачатки и способствовать их росту.
Кущение риса происходит в два этапа: образование пазушной почки в каждой пазухе листа и последующее ее отрастание 2 . Хотя морфология и гистология 2,3 и некоторых мутантов кущения риса 4 хорошо описаны, молекулярный механизм кущения риса еще предстоит выяснить. Здесь мы сообщаем о выделении и характеристике MONOCULM 1 ( MOC1 ), гена, важного для контроля кущения риса. Мутантные растения moc1 имеют только главный стебель без побегов из-за дефекта формирования почек побегов. MOC1 кодирует предполагаемый ядерный белок семейства GRAS, который экспрессируется в основном в пазушных зачатках и функционирует, чтобы инициировать пазушные зачатки и способствовать их росту.
Это предварительный просмотр содержимого подписки, доступ через ваше учреждение
Соответствующие статьи
Статьи открытого доступа со ссылками на эту статью.
Оптимизация архитектуры метелок риса путем специфического подавления пар лиганд-рецептор
- Тао Го
- , Цзы-Ци Лу
- … Хун-Сюань Линь
Связь с природой
Открытый доступ
24 марта 2023 г.Повышенное содержание CO2 как устойчивый подход к увеличению числа побегов риса и потенциальной урожайности
- Дженнифер М. Слоан
- , Аззами Адам Мухамад Муджаб
- … Эндрю Дж. Флеминг
Рис
Открытый доступ
22 марта 2023 г.Может ли дикий многолетний корневищный вид риса Oryza longistaminata быть кандидатом на одомашнивание De Novo?
- Шуай Тонг
- , Мотоюки Асикари
- … Оле Педерсен
Рис
Открытый доступ
16 марта 2023 г.
Варианты доступа
Подпишитесь на этот журнал
Получите 51 печатный выпуск и доступ в Интернете
199,00 € в год
всего 3,90 € за выпуск
Узнать больше
Просто взять напрокат или купить эту статью
3
3 эта статья до тех пор, пока она вам нужна
$39,95
Узнать больше
Цены могут облагаться местными налогами, которые рассчитываются при оформлении заказа
Рисунок 1: Фенотип и дополнение moc1 мутант. Рисунок 2: Молекулярная идентификация MOC1 . Рис. 3: Микроскопические исследования формирования почек риса. Рисунок 4: Продвижение MOC1 в росте побегов и экспрессия OSh2 и OsTB1 в растениях дикого типа и moc1 .
Ссылки
Ли, Ю.-Х. Морфология и анатомия злаковых культур 138–142 (Shanghai Science and Technology Press, Шанхай, 1979)
Google Scholar
Hanada, K. in Science of the Rice Plant Vol. 1 Морфология (ред. Мацуо Т. и Хошикава К.) 222–258 (Исследовательский центр продовольственной и сельскохозяйственной политики, Токио, 1993 г.)
Google Scholar
Ханада, К. в Science of the Rice Plant Vol. 2 Физиология (ред. Мацуо, Т., Кумазава, К., Исии, Р., Исихара, К. и Хирата, Х.) 61–65 (Исследовательский центр продовольственной и сельскохозяйственной политики, Токио, 1995)
Google Scholar
Ивата, Н., Такамуре, И., Ву, Х.-К., Сиддинк, Э.
А. и Рутгер, Дж.Н. Список генов различных признаков (с хромосомой и основной литературой). Рис Жене. Newsl. 12 , 61–93 (1995)Google Scholar
Шумахер К., Шмитт Т., Россберг М., Шмитц Г. и Терес К. Боковой глушитель ( Ls ) ген томата кодирует новый член семейства белков VHIID. Проц. Натл акад. науч. США 96 , 290–295 (1999)
Статья
ОБЪЯВЛЕНИЯ
КАСGoogle Scholar
Пыш, Л. Д., Высоцкая-Диллер, Дж. В., Камиллери, К., Бушез, Д. и Бенфей, П. Н. Семейство генов GRAS в Arabidopsis : характеристика последовательности и базовый анализ экспрессии SCARECROW-LIKE ген. Завод Ж. 18 , 111–119 (1999)
Артикул
КАСGoogle Scholar
Ди Лауренцио, Л. и др. Ген SCARECROW регулирует асимметричное деление клеток, необходимое для создания радиальной организации корня Arabidopsis .
Ячейка 86 , 423–433 (1996)Артикул
КАСGoogle Scholar
Helariutta, Y. et al. Ген SHORT-ROOT контролирует формирование радиального паттерна корня Arabidopsis посредством радиальной передачи сигналов. Ячейка 101 , 555–567 (2000)
Артикул
КАСGoogle Scholar
Bolle, C., Koncz, C. & Chua, N.H. PAT1, новый член семейства GRAS, участвует в передаче сигнала фитохрома А. Гены Дев. 14 , 1269–1278 (2000)
КАС
пабмед
ПабМед ЦентральныйGoogle Scholar
Пэн, Дж. и др. Ген Arabidopsis GAI определяет сигнальный путь, который отрицательно регулирует реакцию гиббереллина. Гены Дев. 11 , 3194–3205 (1997)
Артикул
КАСGoogle Scholar
Сильверстоун, А.
Л., Чампальо, К. Н. и Сан, Т. 9Ген 0137 Arabidopsis RGA кодирует регулятор транскрипции, подавляющий путь передачи сигнала гиббереллином. Растительная клетка 10 , 155–169 (1998)Статья
КАСGoogle Scholar
Itoh, H., Ueguchi-Tanaka, M., Sato, Y., Ashikari, M. & Matsuoka, M. Сигнальный путь гиббереллина регулируется появлением и исчезновением SLENDER RICE1 в ядрах. Растительная клетка 14 , 57–70 (2002)
Статья
КАСGoogle Scholar
Peng, J. et al. Гены «зеленой революции» кодируют мутантные модуляторы ответа гиббереллина. Природа 400 , 256–261 (1999)
Статья
ОБЪЯВЛЕНИЯ
КАСGoogle Scholar
Richards, D. E., Peng, J. & Harberd, N. P. GRAS растений и STAT многоклеточных животных: одно семейство? Bioessays 22 , 573–577 (2000)
Статья
КАСGoogle Scholar
Smith, H.
M., Hicks, G.R. & Raikhel, N.V. Импортин α из Arabidopsis thaliana представляет собой ядерный рецептор импорта, который распознает три класса сигналов импорта. Завод физиол. 114 , 411–417 (1997)Статья
КАСGoogle Scholar
Stirnberg, P., van De Sande, K. & Leyser, H.M. MAX1 и MAX2 боковое ветвление контрольного побега у Arabidopsis . Разработка 129 , 1131–1141 (2002)
CAS
пабмедGoogle Scholar
Катаяма Т. Аналитические исследования кущения риса-сырца. Дж. Имп. Агр. Эксп. Stn Jpn 1 , 327–374 (1931)
Google Scholar
Yan, J.-Q., Zhu, J., He, C.-X., Benmoussa, M. & Wu, P. Количественный анализ локусов признаков поведения числа побегов у риса в процессе развития ( Oryza сатива л.
). Теор. заявл. Жене. 97 , 267–274 (1998)Статья
КАСGoogle Scholar
Сато Ю. и др. Ген гомеобокса риса, ОШ2 , экспрессируется перед дифференцировкой органов в специфической области во время раннего эмбриогенеза. Проц. Натл акад. науч. США 93 , 8117–8122 (1996)
Артикул
ОБЪЯВЛЕНИЯ
КАСGoogle Scholar
Добли, Дж., Стек, А. и Хаббард, Л. Эволюция апикального доминирования у кукурузы. Природа 386 , 485–488 (1997)
Статья
ОБЪЯВЛЕНИЯ
КАСGoogle Scholar
Люкенс, Л. и Добли, Дж. Молекулярная эволюция гена теосинте разветвленного среди кукурузы и родственных трав. Мол. биол. Эвол. 18 , 627–638 (2001)
Статья
КАСGoogle Scholar
Hubbard, L.
, McSteen, P., Doebley, J. & Hake, S. Характер экспрессии и мутантный фенотип teosinte branched 1 коррелируют с подавлением роста у кукурузы и teosinte. Генетика 162 , 1927–1935 (2002)CAS
пабмед
ПабМед ЦентральныйGoogle Scholar
Schmitz, G. et al. Ген томата Blind кодирует фактор транскрипции MYB, контролирующий образование латеральных меристем. Проц. Натл акад. науч. США 99 , 1064–1069 (2002)
Статья
ОБЪЯВЛЕНИЯ
КАСGoogle Scholar
Otsuga, D., DeGuzman, B., Prigge, M.J., Drews, G.N. & Clark, S.E. REVOLUTA регулирует инициацию меристемы в латеральных положениях. Завод Ж. 25 , 223–236 (2001)
Артикул
КАСGoogle Scholar
Harushima, Y.
et al. Карта генетического сцепления риса высокой плотности с 2275 маркерами с использованием одной популяции F2. Генетика 148 , 479–494 (1998)КАС
пабмед
ПабМед ЦентральныйGoogle Scholar
Hiei, Y., Ohta, S., Komari, T. & Kumashiro, T. Эффективная трансформация риса ( Oryza sativa L.), опосредованная Agrobacterium , и анализ последовательности границ T- ДНК. Завод Ж. 6 , 271–282 (1994)
Артикул
КАСGoogle Scholar
Нива Ю., Хирано Т., Йошимото К., Симидзу М. и Кобаяши Х. Неинвазивное количественное определение и применение нетоксичного зеленого флуоресцентного белка типа S65T в живых растениях. Завод Ж. 18 , 455–463 (1999)
Артикул
КАСGoogle Scholar
Фоберт, П.
Р., Коэн, Э. С., Мерфи, Г. Дж. и Дунан, Дж. Х. Закономерности клеточного деления, выявленные путем регуляции транскрипции генов во время клеточного цикла у растений. EMBO J. 13 , 616–624 (1994)Статья
КАСGoogle Scholar
Hu, Y.-X., Bao, F. & Li, J. Стимулирующее действие брассиностероидов на клеточное деление включает особый путь индукции CycD3 у Arabidopsis . Завод Ж. 24 , 693–701 (2000)
Артикул
КАСGoogle Scholar
 А. и Рутгер, Дж.Н. Список генов различных признаков (с хромосомой и основной литературой). Рис Жене. Newsl. 12 , 61–93 (1995)
А. и Рутгер, Дж.Н. Список генов различных признаков (с хромосомой и основной литературой). Рис Жене. Newsl. 12 , 61–93 (1995)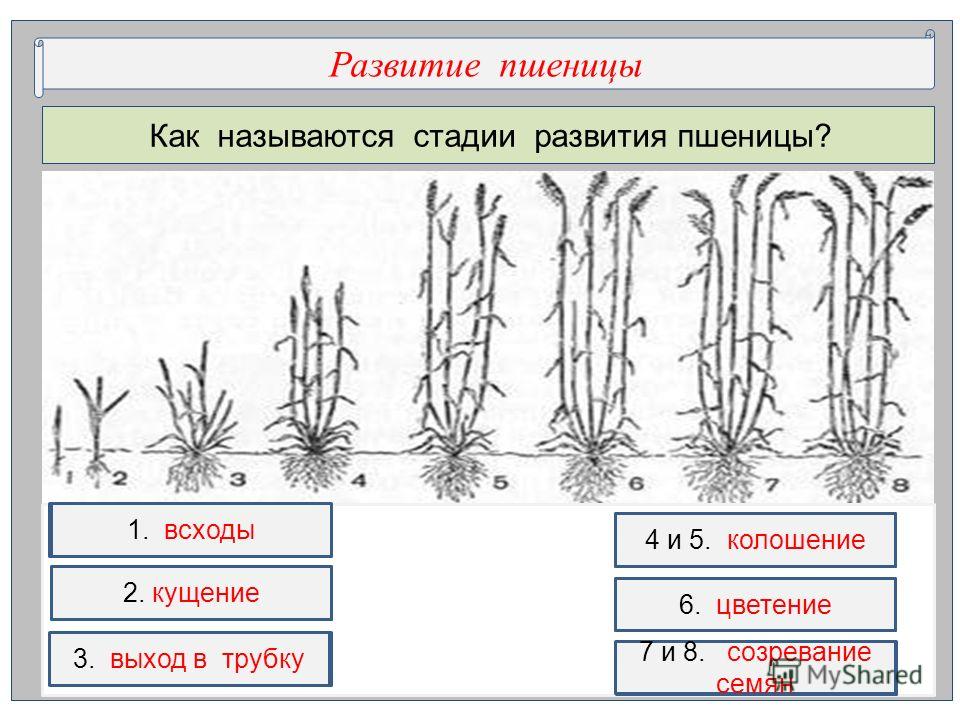 Ячейка 86 , 423–433 (1996)
Ячейка 86 , 423–433 (1996) Л., Чампальо, К. Н. и Сан, Т. 9Ген 0137 Arabidopsis RGA кодирует регулятор транскрипции, подавляющий путь передачи сигнала гиббереллином. Растительная клетка 10 , 155–169 (1998)
Л., Чампальо, К. Н. и Сан, Т. 9Ген 0137 Arabidopsis RGA кодирует регулятор транскрипции, подавляющий путь передачи сигнала гиббереллином. Растительная клетка 10 , 155–169 (1998) M., Hicks, G.R. & Raikhel, N.V. Импортин α из Arabidopsis thaliana представляет собой ядерный рецептор импорта, который распознает три класса сигналов импорта. Завод физиол. 114 , 411–417 (1997)
M., Hicks, G.R. & Raikhel, N.V. Импортин α из Arabidopsis thaliana представляет собой ядерный рецептор импорта, который распознает три класса сигналов импорта. Завод физиол. 114 , 411–417 (1997)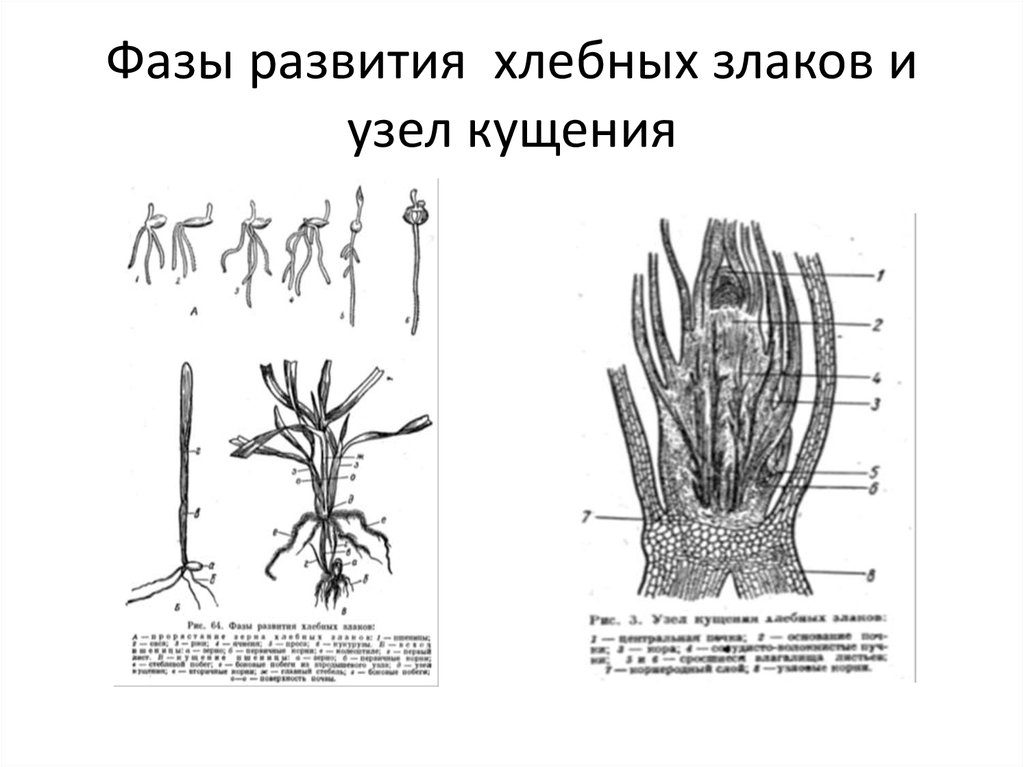 ). Теор. заявл. Жене. 97 , 267–274 (1998)
). Теор. заявл. Жене. 97 , 267–274 (1998) , McSteen, P., Doebley, J. & Hake, S. Характер экспрессии и мутантный фенотип teosinte branched 1 коррелируют с подавлением роста у кукурузы и teosinte. Генетика 162 , 1927–1935 (2002)
, McSteen, P., Doebley, J. & Hake, S. Характер экспрессии и мутантный фенотип teosinte branched 1 коррелируют с подавлением роста у кукурузы и teosinte. Генетика 162 , 1927–1935 (2002) et al. Карта генетического сцепления риса высокой плотности с 2275 маркерами с использованием одной популяции F2. Генетика 148 , 479–494 (1998)
et al. Карта генетического сцепления риса высокой плотности с 2275 маркерами с использованием одной популяции F2. Генетика 148 , 479–494 (1998)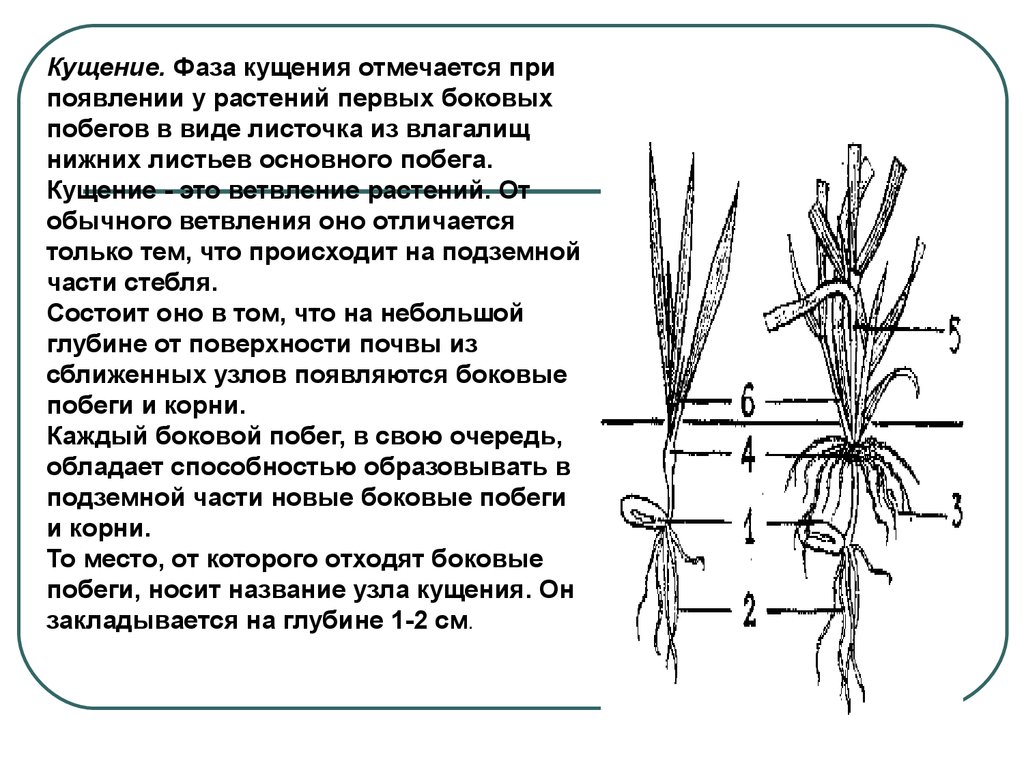 Р., Коэн, Э. С., Мерфи, Г. Дж. и Дунан, Дж. Х. Закономерности клеточного деления, выявленные путем регуляции транскрипции генов во время клеточного цикла у растений. EMBO J. 13 , 616–624 (1994)
Р., Коэн, Э. С., Мерфи, Г. Дж. и Дунан, Дж. Х. Закономерности клеточного деления, выявленные путем регуляции транскрипции генов во время клеточного цикла у растений. EMBO J. 13 , 616–624 (1994)Скачать ссылки
Благодарности
Мы благодарим J. Zuo, N.-H. Чуа и X.-W. Дэну за критические комментарии к рукописи, Б. Чжану за помощь в использовании микроскопа, И. Такамуре за предоставление мутантов rcn1–rcn5, Японскому банку ДНК MAFF за предоставление зондов RFLP и клонов YAC, Геномному институту Университета Клемсона за предоставление Nipponbare Фильтры и клоны библиотеки BAC, Y. Niwa за предоставление конструкции CaMV35SΩ-sGFP (S65T)-NOS-3′ и M. Matsuoka за клон кДНК OSh2 . Ф. Х. — приглашенный ученый из Японского международного исследовательского центра сельскохозяйственных наук, Цукуба, Япония. Эта работа была поддержана грантами Министерства науки и технологий Китая, Национального фонда естественных наук Китая и Китайской академии наук.
Niwa за предоставление конструкции CaMV35SΩ-sGFP (S65T)-NOS-3′ и M. Matsuoka за клон кДНК OSh2 . Ф. Х. — приглашенный ученый из Японского международного исследовательского центра сельскохозяйственных наук, Цукуба, Япония. Эта работа была поддержана грантами Министерства науки и технологий Китая, Национального фонда естественных наук Китая и Китайской академии наук.
Информация об авторе
Примечания автора
Xueyong Li, Qian Qian и Bin Han: Эти авторы в равной степени внесли свой вклад в эту работу , 100101, Пекин, Китай
Сюеюн Ли, Чжиминг Фу, Юнхун Ван, Гуошэн Сюн, Сяоцюнь Ван, Синьфан Лю и Цзяян Ли
Китайский национальный научно-исследовательский институт риса, Китайская академия сельскохозяйственных наук, Ханчжоу, Чжэцзян, 6, Китай
Qian Qian, Dali Zeng, Sheng Teng и Fujimoto Hiroshi
Китайский сельскохозяйственный университет, 100094, Пекин, Китай
Ming Yuan
Институт физиологии растений и экологии, Шанхай0 Китай
Да Луо
Национальный центр исследований генов Китайской академии наук, Шанхай, 200233, Китай
Бин Хань
Авторы
- Xueyong Li
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Qian Qian
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Zhiming Fu
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Yonghong Wang
Посмотреть публикации автора
Вы также можете искать этого автора в
PubMed Google Scholar - Guosheng Xiong
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Dali Zeng
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Xiaoqun Wang
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Академия - Xinfang Liu
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Sheng Teng
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Fujimoto Hiroshi
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Ming Yuan
Посмотреть публикации автора
Вы также можете искать этого автора в
PubMed Google Scholar - Da Luo
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Bin Han
Просмотр публикаций автора
Вы также можете искать этого автора в
PubMed Google Scholar - Jiayang Li
Посмотреть публикации автора
Вы также можете искать этого автора в
PubMed Google Scholar
Автор, ответственный за переписку
Цзяян Ли.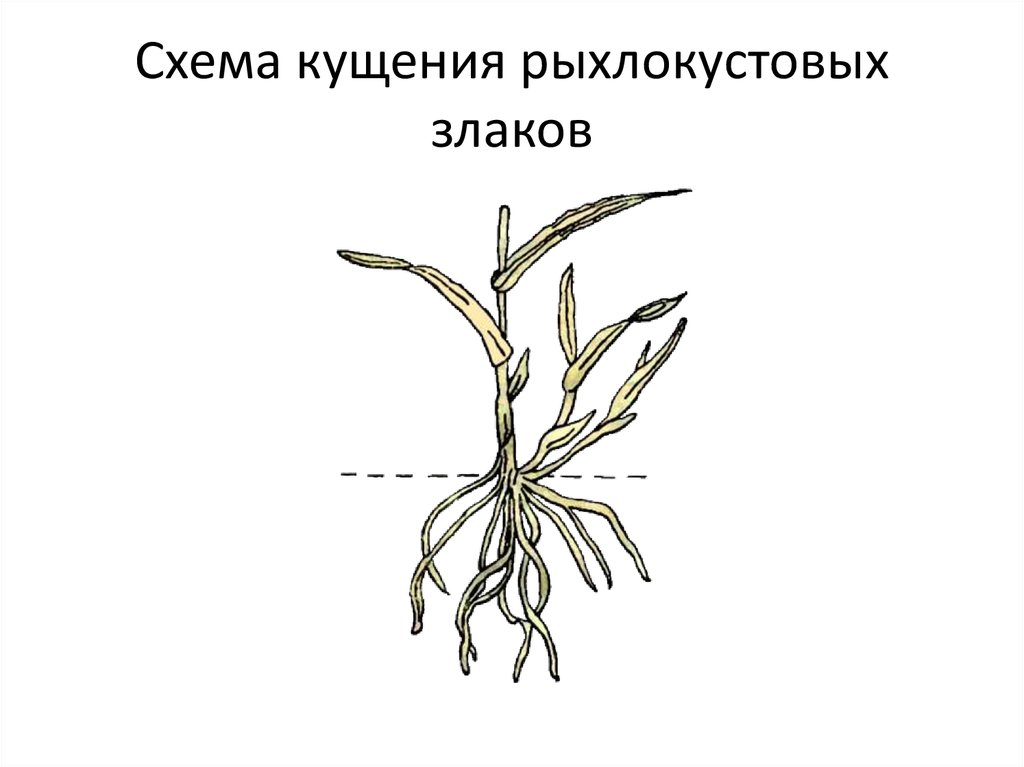
Декларация этики
Конкурирующие интересы
Авторы заявляют, что у них нет конкурирующих финансовых интересов.
Дополнительная информация
Дополнительные таблицы 1 и 2 (DOC 38 kb)
Дополнительный рисунок 1 (PDF 38 kb)
Дополнительный рисунок 2 (PDF, 37 КБ)
Права и разрешения
Перепечатка и разрешения
Об этой статье
Повышенное содержание CO2 как устойчивый подход к увеличению числа побегов риса и потенциальной урожайности
- Дженнифер М. Слоан
- Аззами Адам Мухамад Муджаб
- Эндрю Дж. Флеминг
Рис (2023)
Может ли дикий многолетний корневищный вид риса Oryza longistaminata быть кандидатом на одомашнивание De Novo?
- Шуай Тонг
- Мотоюки Асикари
- Оле Педерсен
Рис (2023)
Молекулярная характеристика показывает, что OsSAPK3 улучшает засухоустойчивость и урожайность риса.

- Дэнцзи Лу
- Супин Лу
- Сяоянь Ян
BMC Биология растений (2023)
Оптимизация архитектуры метелок риса путем специфического подавления пар лиганд-рецептор
- Тао Го
- Цзы-Ци Лу
- Хун-Суан Линь
Nature Communications (2023)
Tiller Number1 кодирует белок анкириновых повторов, который контролирует кущение мягкой пшеницы.